

{kind=link}
{kind=link}
{kind=link}
{kind=link}
瘤突苍耳入侵的种子生态学机制
[曲波1  , 荀挚峰
, 荀挚峰2 , 许玉凤1 ]
, 荀挚峰]
|
|


第一作者:曲波(1972-),女,辽宁朝阳人,副教授,博士,主要从事入侵生物学与生物多样性。E-mail:[email protected]
外来植物瘤突苍耳( Xanthium strumarium)已成为我国东北地区危害较重的入侵植物之一,为明确其是否能与本地同属近缘种苍耳 ( X. sibiricum)杂交,对两种植物及互交后的果实形态、种子的萌发特性进行了比较研究。结果表明,瘤突苍耳自交果实与互交( X. strumarium×X. sibiricum)果实的正面观面积、周长、长和宽、果实重量、萌发率及萌发速率均显著高于苍耳自交果实与互交( X. sibiricum×X. strumarium)果实( P<0.05)。瘤突苍耳互交(Xst♀×Xsi♂)果实长度、周长和刺长均大于瘤突苍耳自交果实( P<0.05),二者果实面积和宽度、果实百粒重、种子萌发率及萌发速率都无显著差异( P>0.05);苍耳自交果实和苍耳互交(Xsi♀×Xst♂)果实仅在果实宽度和种子萌发速率上有明显差异,苍耳互交(Xsi♀×Xst♂)果实宽度高于苍耳自交果实( P<0.05),种子萌发速率低于苍耳自交种子( P<0.05)。由此可见,瘤突苍耳与苍耳及互交后的果实特性具有明显差别,瘤突苍耳杂交后果实明显大于其母本瘤突苍耳,苍耳杂交后与其母本苍耳形态差异不大,表明瘤突苍耳杂交后生活能力增强,较其母本及本地种苍耳能更早占据生长空间与资源,从而提高其入侵能力。
Exotic plants Xanthium strumarium in Northeast China has become one of the most serious invasive plants. In order to determine whether it can hybridize with native plants X. sibiricum. Acomparison of invasive plants X. strumarium, native plants X. sibiricumandthe reciprocal crossing fruits has been studied and the differences between fruit and germination characteristics have been discussed. The results showed that the front view area, perimeter, length and width, fruits weight, germination rate and speed of the reciprocal crossing ( X. strumarium×X. sibiricum) and the inbred X. strumarium fruits were significantly higher than those of the reciprocal crossing ( X. sibiricum×X. strumarium) and the inbred X. sibiricum ( P<0.05). The perimeter, length and length of thorns of the reciprocal crossing fruits of X. strumarium(Xst♀×Xsi♂)were significantly longer than those of inbred X. strumarium fruits ( P<0.05). Both the area, width, fruits weight, germination rate difference between these two fruits were not significant. The width and germination rate of reciprocal crossing fruits of X. sibiricum (Xsi♀×Xst♂) were significantly different with those of inbred X. sibiricum fruit ( P<0.05). The width of the reciprocal crossing fruits of X. sibiricum (Xsi♀×Xst♂)was significantly higher than that of inbred X. sibiricum fruit whereas the germination rate was opposite. Therefore, the difference of fruit morphological characteristics between these two plants and their hybrid was significantly different. The reciprocal crossing fruits of X. strumarium(Xst♀×Xsi♂)was significantly higher than that of the inbred X. strumarium. The difference between the reciprocal crossing seeds of X. sibiricum (Xsi♀×Xst♂) and the inbred X. sibiricum seeds was not significant. The results suggested hybrid improve X. strumarium quality which may occupy space and resources more earlier than X. strumarium and X. sibiricum to increase the chance to invase.
外来生物入侵已成为全球性问题之一。入侵生物破坏当地生物多样性, 打破生态平衡并加速物种灭绝, 有的甚至威胁人类健康, 对当地生态、经济、社会等产生严重影响[1, 2, 3]。与本地物种相比, 外来入侵种具有较高的生态适应性和协同进化能力, 生长繁殖迅速, 可以通过传播疾病、竞争、捕食等方式, 影响本地物种的生态系统功能[4, 5, 6]。研究发现, 入侵植物可通过与本地同属近缘种植物杂交, 改变本地物种的遗传多样性和提高入侵植物对环境的适应性、竞争能力, 进而影响本地物种生存[7, 8, 9, 10, 11, 12, 13]。
植物种子作为植物界演化的最高阶段, 是种子植物生活史中一个重要的生育期, 有更强的抗逆性[14]。而萌发又是种子生活史中一个重要阶段, 直接关系到种群的生存和发展, 同时也是植物适应不同生态环境进而维持种群繁衍而形成的一种进化特性[15]。所以, 植物种子特性与植物生长密切相关, 探究植物种子特性与入侵性的关系, 可为研究外来植物入侵提供理论依据。
瘤突苍耳(Xanthium strumarium), 别名意大利苍耳, 是一种具有二形性种子的一年生菊科植物, 原产于北美洲, 随进口农副产品或包装物携带传入我国, 是一种重要的危险性植物, 已被我国列入《中华人民共和国进境植物检疫性有害生物名录》[16, 17, 18]。瘤突苍耳果实产量高, 对环境的适应能力强, 尤其是在湿润地带能快速形成优势种群, 其在东北地区已成为危害较重的入侵植物之一, 给农作物生产和生态环境带来严重危害[19, 20, 21, 22]。沈阳农业大学生物科学技术学院通过3年野外观察, 发现瘤突苍耳与本地同属近缘种植物苍耳(X. sibiricum)花期一致。那么入侵种瘤突苍耳与本地种苍耳是否能杂交?若能杂交, 杂交果实和种子特性有何变化?本研究以入侵植物瘤突苍耳和本地近缘种苍耳及二者互交后代为材料, 研究互交前后果实和种子变化情况, 探讨杂交是否能促进瘤突苍耳的入侵性, 为瘤突苍耳的防治提供依据。
2012年11月于辽宁省凌海市右卫满族镇小黄村(121° 34'50.24″ E, 41° 04'41.81″ N)河岸山地瘤突苍耳自然成熟植株上采集果实, 在沈阳市东陵区东陵公园(123° 35'06.05″ E, 41° 49'43.13″ N)附近弃耕地苍耳自然成熟植株上采集果实, 均置于通风避光处晾干, 室温保存备用。
杂交试验于2013-2014年在沈阳农业大学北山实验基地进行。在瘤突苍耳与苍耳盛花期时进行自花授粉和杂交授粉, 共4个处理, 每处理80个花序。自交组合两组:瘤突苍耳自交和苍耳自交, 本文分别简称瘤突苍耳和苍耳。互交组两组:瘤突苍耳× 苍耳(Xst♀× Xsi♂), 本文简称瘤突苍耳互交; 苍耳× 瘤突苍耳(Xsi♀× Xst♂), 本文简称苍耳互交。授粉花序分别严格套袋隔离。待花期过后去除杂交袋, 分别采集成熟后的果实, 置于通风避光处晾干, 室温保存备用。
1.3.1 果实形态及重量特征测定 分别取瘤突苍耳和苍耳自交及互交果实各100粒, 1/10 000天平称量其百粒重, 重复5次, 计算平均值。每个处理随机取出30粒果实进行形态测定, 主要指标为正面观面积、周长、长(果实纵轴长度)、宽(果实横轴长度)、刺长等。具体操作方法按照王学治等[23]的方法, 根据与标准对照的像素比例测定果实的正面观面积、周长、刺长、长度、宽度(图1)。
 | 图1 瘤突苍耳果实的形态指标Fig.1 The shape index of the fruit of X. strumarium |
1.3.2 萌发率测定 将去除果苞的苍耳、瘤突苍耳及互交果实的种子放在垫有滤纸培养皿中, 每个培养皿放30粒种子, 保持适当水分, 置于光照培养箱中(温度25 ℃, 光照12 h), 出现胚根即认为已经萌发, 3次重复。因为苍耳与瘤突苍耳都具有二形性种子[16], 为了保持试验条件的一致性, 所用种子均为上位种子。
初始萌发天数是指种子萌发距离萌发试验开始的天数(d); 萌发高峰期是指萌发量最大的时间距萌发试验开始的天数(d)。萌发持续时间是指从萌发试验开始到最后一个种子萌发的总天数(d)。萌发速率是指种子萌发率达到50%时所用的天数(d)。
用SPSS 16.0进行数据处理, 用Duncan法对果实形态数据和萌发数据进行显著性分析, 用Photoshop CS 4和Microsoft Excel作图。
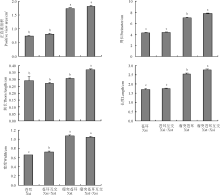
瘤突苍耳与苍耳自交和互交果实形态特征有所不同。通过对各组合果实形态指标测量发现, 瘤突苍耳互交果实正面观面积最大, 约为1.806 cm2, 苍耳自交果实面积最小, 约为0.737 cm2; 瘤突苍耳互交果实周长最大, 约为7.787 cm, 苍耳自交果实周长最小, 约为4.306 cm; 瘤突苍耳互交果实刺长最大, 约为0.368 cm, 苍耳互交果实刺长最小, 约为0.269 cm; 瘤突苍耳互交果实长度最大, 约为2.772 cm, 苍耳自交果实长度最小, 约为1.704 cm; 瘤突苍耳自交果实宽度最大, 约为1.067 cm, 苍耳自交果实宽度最小, 约为0.651 cm(图2)。瘤突苍耳自交果实的正面观面积、周长、长和宽高于苍耳自交果实(P< 0.05), 二者果实的刺长无差异(P> 0.05)。瘤突苍耳互交果实的周长、刺长和长度高于瘤突苍耳自交果实, 二者果实的正面观面积和宽度无差异。苍耳自交果实与互交果实果实正面观面积、周长、刺长和长度都差异不明显(P> 0.05), 仅宽度上苍耳互交果实高于苍耳自交果实。瘤突苍耳互交果实各个指标上都高于苍耳互交果实(P< 0.05)。
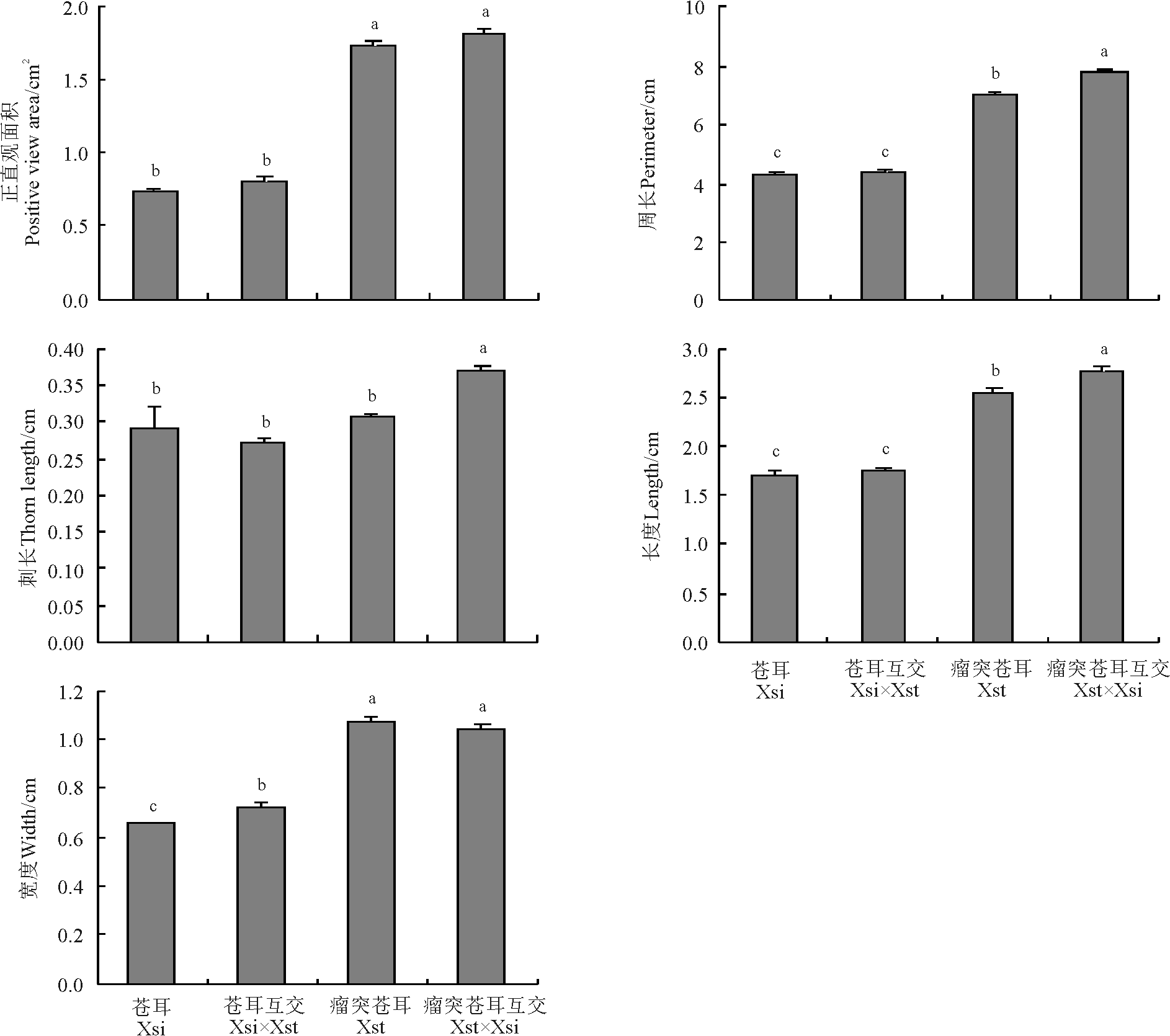 | 图2 瘤突苍耳与苍耳自交果实与互交的果实的形态特征比较注:不同小写字母表示果实之间差异显著(P< 0.05)。下同。Fig.2 Comparison of positive view area, perimeter, thorn length, width and length of fruit of X. strumarium, X. sibiricum and the reciprocal crossing fruitsNote: Different lower case letters indicate significant difference among different fruits at 0.05 level. The same below. |
瘤突苍耳与苍耳的自交和互交果实粒重有所不同。通过称量各组合果实百粒重, 发现瘤突苍耳互交果实百粒重最大, 为38.847 g, 苍耳果实百粒重最小, 为11.739 g(图3), 瘤突苍耳果实和苍耳互交百粒重分别为34.955 g和14.392 g。瘤突苍耳和瘤突苍耳互交果实百粒重显著大于苍耳和苍耳互交(P< 0.05)。瘤突苍耳果实与瘤突苍耳互交果实百粒重无明显差异(P> 0.05), 苍耳果实与苍耳互交果实百粒重也无明显差异(P> 0.05)。
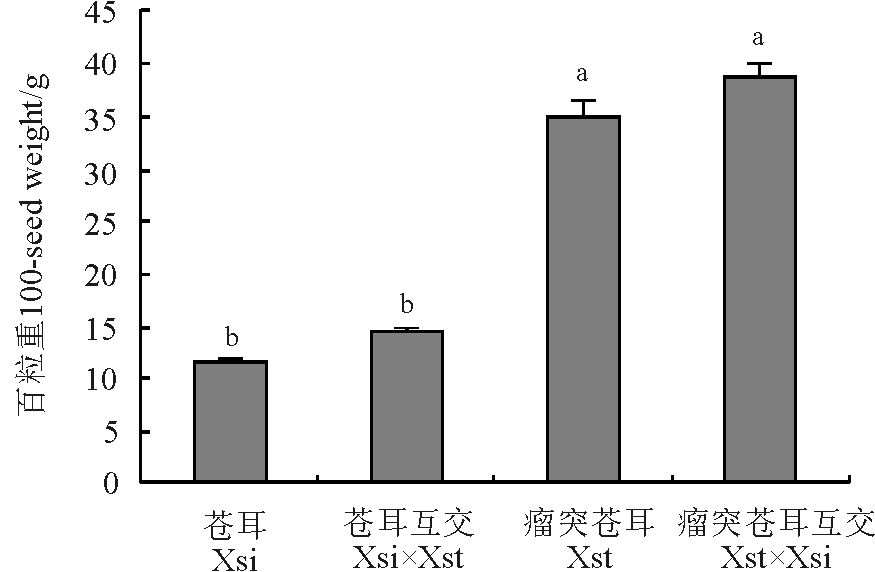 | 图3 瘤突苍耳与苍耳自交及互交果实百粒重Fig.3 Comparison of fruit mass of X. strumarium, X. sibiricum and the reciprocal crossing fruits |
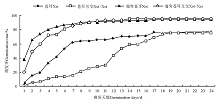
瘤突苍耳种子、瘤突苍耳互交种子和苍耳种子从试验开始的第1天开始萌发, 苍耳互交种子第2天开始萌发。瘤突苍耳种子和瘤突苍耳互交种子的萌发率及萌发速率高于苍耳种子与苍耳互交种子(P< 0.05)。瘤突苍耳种子萌发率达到最终萌发率的50%的时间比瘤突苍耳互交种子早2 d(P> 0.05), 二者萌发率差别不大(P> 0.05); 苍耳种子萌发率达到最终萌发率的50%的时间比苍耳互交种子早7 d(P< 0.05), 且二者萌发率变化也不大(P> 0.05)(图4)。瘤突苍耳种子萌发最快, 萌发高峰为1-7d, 萌发持续17d, 2d时萌发率达到最终萌发率的50%, 最终萌发率为94.43%; 瘤突苍耳与瘤突苍耳互交种子的萌发高峰期为1-7 d, 萌发持续14 d, 3 d时萌发率达到最终萌发率的50%, 最终萌发率为95.57%; 苍耳种子萌发高峰期1-7d, 萌发持续21d, 6d时萌发率达到最终萌发率的50%, 最终萌发率为76.67%; 苍耳互交种子萌发最慢, 萌发高峰期为11-17d, 萌发持续19d, 13 d时萌发率达到最终萌发率的50%, 最终萌发率为75.53%。
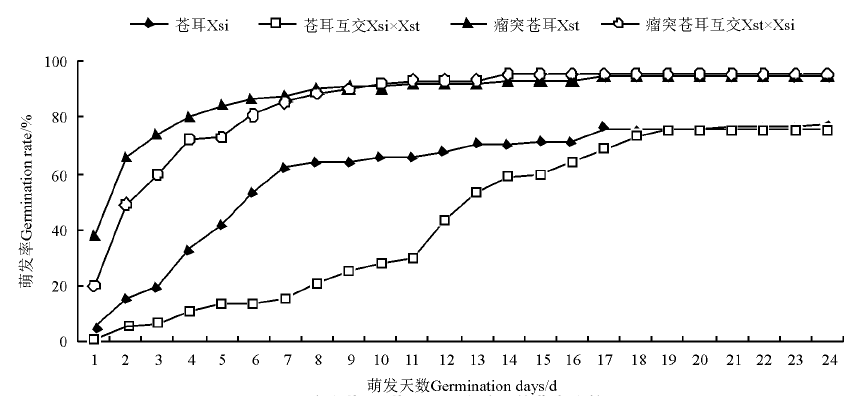 | 图4 瘤突苍耳、苍耳及互交种子的萌发比较Fig.4 Comparison of seed germination of X. strumarium, X. sibiricum and the reciprocal crossing fruits |
果实形态特征不仅表达了一定量较为稳定的遗传信息, 而且与果实的生产、脱落、传播、贮备、萌发与定植等过程密切相关, 从而影响植被的恢复与演替[24]。果实形态特征主要包括种子的重量、大小、形状、表面及其附属物、颜色及光泽等。瘤突苍耳果实与本地苍耳果实形状类似, 但瘤突苍耳果实较苍耳果实较大较重, 单株可结约1 400粒果实, 这表明瘤突苍耳能凭借其结实量迅速占领生境, 从而逃避风险、繁殖后代。大的果实和高的结实量能保证植物有更多的种子或果实进入土壤种子库, 为植物的生存提供了大量种源, 从而有利于植被的更新, 对种群补充后代、稳定发展具有重要作用[25]。
Jurado和Estrada[26]对种子和果实的大小进行研究, 认为小的种子或果实(< 1 mg)代表了风传播类型, 中等大小的种子或果实(1~9.9 mg)是各种传播类型的过渡阶段, 大的种子或果实(> 9.9 mg)主要是动物传播类型。瘤突苍耳和苍耳的单个果实重量均大于9.9 mg, 属于动物传播类型, 且表面具有钩刺, 主要靠重力传播。同时由于现在人类活动频繁, 种子或果实的传播已经突破传统自然力的限制, 瘤突苍耳已被证明能够与北方谷类种子混杂, 增加了传播机会, 这也大大促进了瘤突苍耳入侵性。
植物种子萌发行为是植物生活史繁殖对策研究的重要方面, 植物群落的形成、演替与种子萌发特性有关[27]。其中种子萌发率和萌发速率是衡量生长能力的重要指标[28]。Greipsson和Davy[29]的研究表明, 小种子种群的萌发百分数为大种子种群的50%, 萌发一般所需时间为大种子种群的10倍; Happer等[30]研究也表明, 小种子萌发明显慢于大种子。这是由于大的种子内含有较多的能量物质, 为种子的萌发提供更多的营养物质, 也有助于适应恶劣环境条件[31, 32, 33, 34, 35]。瘤突苍耳种子的萌发率和萌发速率显著高于苍耳种子(P< 0.05), 表明瘤突苍耳较苍耳可以更早获得营养生长和生殖生长的机会, 能更有效地占领当地空间和资源, 使其植株具有更强竞争能力。
瘤突苍耳与苍耳互交后, 瘤突苍耳果实形态特征和苍耳种子的萌发特性发生了明显变化。瘤突苍耳互交果实的周长、刺长和长度大于母本瘤突苍耳果实; 瘤突苍耳的正直观面积、周长、长和宽、重量都高于苍耳果实与苍耳互交果实。可见, 杂交后的瘤突苍耳果实的生活力较母本及杂交苍耳果实有增强趋势; 瘤突苍耳互交种子的萌发速度和萌发率显著高于苍耳自交种子和苍耳互交种子(P< 0.05), 且苍耳互交种子的萌发速度显著低于其母本(P< 0.05)。说明在环境条件适宜时, 杂交后的瘤突苍耳种子较杂交苍耳可以更早获得营养生长和生殖生长的机会, 能更有效地占领当地空间和资源。由此推测, 瘤突苍耳杂交后代生活能力增强, 较其母本及本地种苍耳能更早地占据生长空间和利用资源, 增加其竞争力, 促进其入侵性。
另外, 有遗传研究表明, 果实周长、果长、果宽等形态特征在不同单株间均存在显著或极显著差异, 说明果实形态性状在单株水平上存在丰富的遗传变异[36]。瘤突苍耳与苍耳互交后, 瘤突苍耳互交果实较母本瘤突苍耳的果实形态变化较大, 这可能是由于杂交导致的遗传变异。瘤突苍耳接受苍耳花粉后果实形态变化较多, 苍耳接受瘤突苍耳花粉后果实形态变化不大, 这说明苍耳对瘤突苍耳的影响大于瘤突苍耳对苍耳的影响。这些影响是否有利于提高瘤突苍耳后代的生活力、竞争力和适应性, 是否会促进其入侵有待进一步研究。
The authors have declared that no competing interests exist.
| [1] |
|
| [2] |
|
| [3] |
|
| [4] |
|
| [5] |
|
| [6] |
|
| [7] |
|
| [8] |
|
| [9] |
|
| [10] |
|
| [11] |
|
| [12] |
|
| [13] |
|
| [14] |
|
| [15] |
|
| [16] |
|
| [17] |
|
| [18] |
|
| [19] |
|
| [20] |
|
| [21] |
|
| [22] |
|
| [23] |
|
| [24] |
|
| [25] |
|
| [26] |
|
| [27] |
|
| [28] |
|
| [29] |
|
| [30] |
|
| [31] |
|
| [32] |
|
| [33] |
|
| [34] |
|
| [35] |
|
| [36] |
|