{kind=link}
PEG与低温对法国百里香种子萌发的影响
[赵波1  , 杨秀云
, 杨秀云2 , 沈向群1 ]
, 杨秀云]
|
|


第一作者:赵波(1976-),女,河南沁阳人,讲师,在读博士生,主要从事草类种子与种质资源研究。E-mail:[email protected]
以法国百里香( Thymus vulgaris)为材料,研究了PEG模拟干旱胁迫和低温对种子萌发和幼苗生长的影响,旨在探索进口百里香种子播种育苗的适宜条件。结果表明,温度、干旱及其互作对法国百里香种子萌发和幼苗生长均有显著影响。0~10% PEG-6000水分条件下,8~24 ℃下法国百里香种子均能良好萌发,其中在10~12 ℃时发芽率最高,达79.3%,显著高于其余各处理( P<0.05)。当PEG-6000达15%时显著抑制其萌发,至30% PEG-6000时仅在22~24 ℃下少量萌发。萌发后,当温度为22~24 ℃,根长、茎长均显著高于其他处理。种子和幼苗生长均表现出对PEG模拟干旱胁迫和低温不同的响应。
In order to understand the optimal conditions for germination and seedling growth of import thyme seeds—— Thymus vulgaris, the effects of PEG-simulated drought stress and low temperature on seed germination and seedling growth were measured. The analysis of two-way ANOVA showed that temperature, drought and their interaction significantly affected ( P<0.05) the seed germination and seedling growth. The treatments with the temperature ranged from 8 ℃ to 24 ℃ and PEG-6000 ranged from 0 to 10% were optimal for seeds germination. The treatment with the temperature at 10 ℃ to 12 ℃ had the highest germination rate of 79.3%, which had significant difference from the other treatments ( P<0.05). The treatments with 15% PEG-6000 could significantly inhibit ( P<0.05) seed germination. Under the treatments with 30% PEG-6000, seeds only could germinate at 22 ℃ to 24 ℃. The root and shoot length under treatments with the temperature at 22 ℃ to 24 ℃ were significantly higher than the other treatments. Seeds and seedling growth had different response to PEG-simulated drought stress and cold.
百里香属(Thymus)植物花期长, 花色艳丽, 耐旱抗寒, 全株散发芳香气味, 是一种优良的芳香观赏草, 宜栽植在岩石园、花境、花坛、香料园或向阳处等, 在城市园林绿化中具有广阔的应用前景[1]。百里香属植物常呈地毯状连片生长, 多生于山地、山谷、山沟及路旁。我国的百里香属植物多分布于西北、华北和东北地区, 在海拔1 100~3 600 m的溪旁、石山地、黄土丘陵和沙地上较常见[2, 3]。百里香属植物用途广泛, 在香料、调料、医药、园林绿化等行业均有应用。其新鲜枝叶可生食, 做厨房料理之香料; 全株泡水, 具防腐抗菌之效用; 炮制加工, 是增强免疫之良药[1, 4]。
随着百里香栽培面积日益扩大, 其栽培技术已成为重要的研究内容, 而百里香种子萌发和幼苗生长阶段是其栽培群体构建的关键时期。研究表明, 温度、光照、覆土厚度、播种方式、赤霉素浸种等因素对百里香种子萌发和幼苗生长有一定的影响[5, 6, 7]。目前, 试验材料以百里香野生种为主, 而其花期长, 种子成熟不一致, 落粒性强, 不易大量采收种子, 且种子脱萼困难, 不去萼的种子发芽率极低。进口百里香种子在我国百里香栽培生产中用量增大, 而百里香种子小, 千粒重约为0.27 g, 价格昂贵, 种子萌发过程中对水分和温度等环境因子需求敏感, 种子播种存在发芽率和成苗率都很低的问题。因此, 摸清进口百里香种子的萌发特点对其生产栽培具有指导意义。法国百里香(T. vulgaris)香气浓郁, 应用范围广, 表现突出, 是进口百里香的重要代表[8, 9, 10]。本研究以法国百里香为研究对象, 探讨法国百里香种子萌发和幼苗生长对干旱、低温及双重胁迫的响应机制, 旨在为进口百里香的种子播种育苗及园林推广应用提供理论基础和实践指导。
试验选用北京南无科贸有限责任公司花仙子品牌的法国百里香种子, 生产日期均为2013年10月。选取大小一致、颗粒饱满、无病虫害的饱满种子为试验材料, 供试验用的法国百里香幼苗是由种子萌发培养而得。
本试验采用聚乙二醇-6000(PEG-6000)模拟干旱, 分别设置0(对照)、10%、15%、30%共4个处理梯度, 分别置于8~10 ℃、12~14 ℃、22~24 ℃ 3个温度梯度下, 每个处理设3次重复, 每个重复50粒种子。采用纸上发芽方法, 种子均匀置于铺有滤纸的培养皿(90 mm)中, 试验时用不同浓度的PEG-6000溶液浸润滤纸, 然后把培养皿置于不同温度的光照培养箱中[11]。每24 h向滤纸加PEG-6000溶液数滴, 保持滤纸湿润为宜, 每2 d更换一次滤纸, 试验共进行27 d。
以根的长度≥ 种子直径的2倍作为发芽标准, 从种子置床开始每日记录发芽个数。将3个重复中第1粒种子发芽日期作为该处理的发芽开始期, 以种子发芽率没有变化之日作为发芽结束期。试验中统计种子的发芽个数及发芽时间, 发芽结束后计算发芽率、发芽势、发芽指数(GI)[12]。
发芽势=规定日期内发芽种子数/供试种子数× 100%。
发芽率=最终发芽种子数/供试种子数× 100%。
发芽指数(GI)=∑ Gt/Dt (Gt为在第t日的发芽数, Dt为发芽天数)。
种子发芽结束后时, 测量培养皿中所有正常生长的幼苗的生长指标。取样、照相、测量, 用CAD软件的光栅图像参照功能将图片缩放到实际大小, 测量幼苗的根长(mm)、茎长(mm)以及叶面积(mm2), 取平均值进行对比。
利用Microsoft Excel 2003进行数据录入、计算、作图。采用SPSS 17.7统计软件中AVONA对百里香种子萌发和幼苗生长数据进行方差分析, LSD多重比较法分析处理间差异, 显著水平为0.05。
温度是植物种子萌发和出苗的基本条件之一[13, 14], 低温会延迟法国百里香种子萌发的初始时间。法国百里香种子在22~24 ℃条件下第4天开始萌发, 在12~14 ℃条件下, 第5天开始发芽, 当温度降到8~10 ℃时, 第11天种子才开始发芽(图1)。
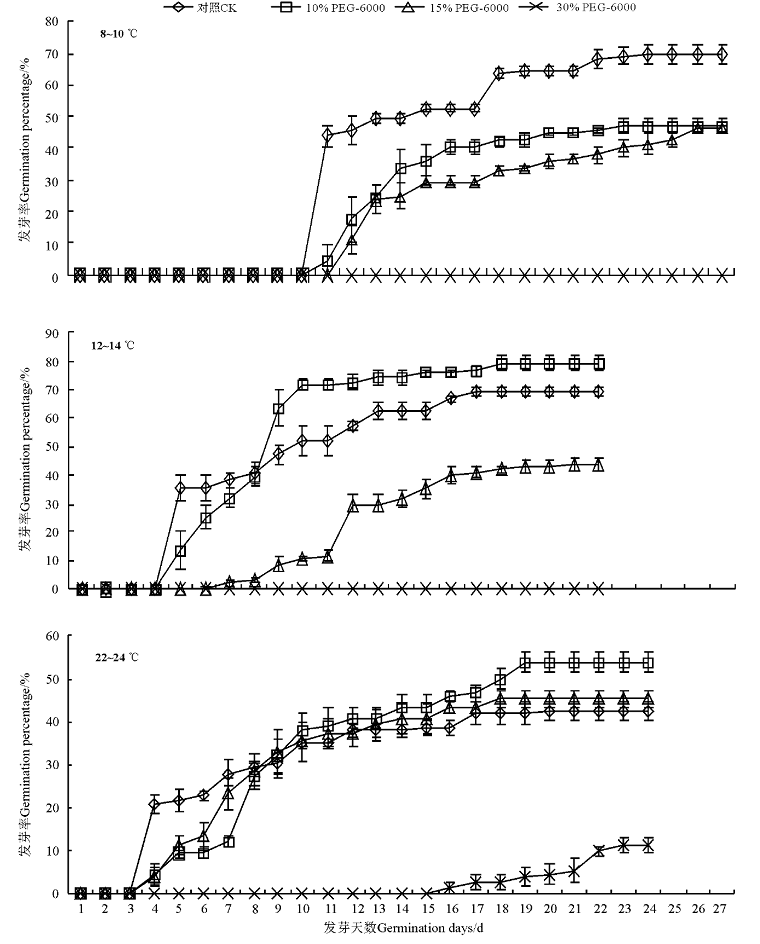 | 图1 不同温度下模拟干旱胁迫对法国百里香种子发芽率的影响Fig.1 Effects of PEG-simulated drought stress on the germination rate of Thymus vulgaris seeds under different temperatures |
PEG-6000干旱处理影响了种子发芽起始时间, 这种影响和PEG-6000处理浓度、发芽温度条件有关。在3个温度条件下, 10% PEG-6000处理对法国百里香种子开始发芽时间无影响; 在22~24 ℃条件下, 15% PEG-6000处理对起始发芽时间无影响, 而温度为12~14 ℃、8~12 ℃时, 起始发芽时间延迟, 分别为第8天和第12天。30% PEG-6000处理对法国百里香种子发芽影响较大, 在8~10 ℃、12~14 ℃时, 种子不能发芽; 在22~24 ℃时, 种子在第13天才开始发芽(图1)。
发芽率反映种子在适宜条件下的萌发能力, 发芽势重在反映种子的活力和品质[15, 16]。未经PEG处理的种子, 在8~10 ℃, 12~14 ℃, 22~24 ℃时, 种子发芽率分别为69.7%, 69.3%和43.3%, 即低温下种子发芽率显著升高(P< 0.05)。30% PEG-6000处理严重影响种子发芽率, 在8~10 ℃, 12~14 ℃时, 发芽率为0, 在22~24 ℃时, 为11.3%; 15%PEG处理后, 发芽率约45%, 3个温度处理无显著差异(P> 0.05); 10% PEG-6000处理后, 在8~10 ℃条件下, 发芽率为47.3%, 和15% PEG-6000各处理间差异不显著, 而在12~14 ℃时, 发芽率为79.3%, 显著(P< 0.05)高于各处理和对照(图1、表1)。这说明PEG-6000处理总体上抑制了法国百里香种子发芽, 尤其是30% PEG-6000处理时, 但是, 在12~14 ℃, 10% PEG-6000处理种子可显著促进法国百里香种子的萌发。
| 表1 干旱与低温对法国百里香种子萌发的影响 Table 1 Effects of drought stress and temperature on the seed germination of Thymus vulgaris |
8~24 ℃范围内, 温度变化对法国百里香幼苗的叶面积、根茎比影响不显著(P> 0.05), 而对其根长与茎长影响显著(P< 0.05)(表2, 表3)。3个温度处理间茎长差异显著, 22~24 ℃时, 茎长最长, 为6.0 mm, 8~10 ℃时次之, 12~14 ℃时最小; 而22~24 ℃时, 幼苗的根长显著高于8~10 ℃和12~14 ℃时的, 后两者之间无显著差异。综合各指标变化效应说明, 22~24 ℃更有利于法国百里香幼苗根长、茎长的生长。
| 表2 干旱与低温对法国百里香幼苗生长的影响 Table 2 Effects of drought stress and temperature on seedling of Thymus vulgaris |
| 表3 温度、干旱对法国百里香幼苗生长的双因素方差分析F值 Table 3 Effects of temperature and drought stress on seedling of Thymus vulgaris by Two-way AVONA |
除了30% PEG-6000处理外, 其他浓度对法国百里香幼苗叶面积、根茎比无显著影响(P> 0.05), 而随着干旱胁迫程度的增加, 法国百里香根长和茎长均呈下降趋势。10% PEG-6000处理与对照的茎长、根长差异不显著, 而15% PEG-6000、30% PEG-6000处理均使根长、茎长均显著减小(P< 0.05)。说明重度干旱胁迫对法国百里香幼苗生长有明显抑制作用, 但轻度胁迫对幼苗生长影响不显著。方差分析结果显示, 干旱以及温度与干旱互作对法国百里香幼苗生长各项指标均有显著影响, 其中对叶面积、根长和根茎比的影响极显著(P< 0.01)。
温度对种子吸胀、萌芽、胚根伸长、胚芽伸长至幼苗形成的整个过程均有重要作用, 它直接影响种子萌发或通过间接破除种子休眠影响种子萌发。目前, 关于干旱与低温胁迫对植物种子萌发的影响研究很多, 比如对番茄(Lycopersicon esculentum)[17]、木蓝(Indigofera tinctoria)[18]和绵枣儿(Scilla scilloides)[19]种子的研究表明, 干旱与低温的作用严重抑制了种子的萌发, 在一定温度范围内, 随温度的升高, 种子萌发进程加快。分析桔梗(Platycodon grandiflorus)[20]、醉马草(Achnatherum inebrians)[21]、决明(Cassia obtusifolia)[22]种子萌发对干旱、低温胁迫的响应发现, 干旱与低温的作用严重抑制了幼苗的生长。本研究表明, 法国百里香种子在12~14 ℃、PEG-6000浓度为10%时发芽率最高, 这可能与低温打破其休眠, 激活其种子萌动进程有关。百里香种子细小, 能供给种子萌发的贮藏物质数量十分有限, 百里香种子萌发过程中, 当其吸水后, 种皮外层的胶状物吸水变软包裹着整个百里香种子, 当温度过高时, 种子易受霉菌侵袭, 不易发芽, 适当的低温处理, 破除法国百里香种子的休眠, 同时种皮外胶状物与低浓度PEG-6000作用, 保持水分缓慢而持续地被种子吸收, 从而使其发芽率较高。
种子萌发和幼苗生长是一个复杂的植物生理生化、物质代谢过程, 受其内部或外部生长物质及环境因子的调控作用, 直接或者间接影响植物的生长和发育[23]。法国百里香的胚根、胚芽以及叶在PEG-6000浓度为30%的时候基本无法生长; 在浓度为15%的情况下, 幼苗生长不整齐, 随着温度的变化呈现出了显著差异; 在浓度为10%的情况下, 幼苗可以正常生长, 随着温度的上升突显其生长优势。不同温度处理下的生长情况表现出显著的差异性, 当温度上升为22~24 ℃时, 幼苗的生长明显优于其他处理。本研究显示, 轻度、中度干旱胁迫对法国百里香根、茎伸长影响较小。其原因可能与百里香种子的结构特征有关。在本研究中, 不论采收的野生百里香种子还是购买的进口百里香种子, 在高倍体视显微镜下没有发现其种子表皮有什么特殊的物质; 当种子遇水吸胀后, 首先是种皮外围一层亲水胶状物胀大变厚, 种子变的很滑, 然后才是种皮吸胀、破裂, 胚发芽。低浓度、少量的PEG-6000可能打破了严密的亲水胶状物的保护, 加快了吸胀进程, 促进了种子萌发, 而高浓度的PEG-6000在打破了严密的亲水胶状物的保护后, 对胚的萌发起到了抑制作用。可见, 适当的低温和PEG-6000处理的条件可促进百里香种子萌发。
PEG作为一种高分子渗透剂, 可以人为地限制水分进入种子的速率从而控制种子的吸水速率和发芽进程[24]。本研究通过PEG-6000模拟干旱胁迫在不同的低温条件下, 研究法国百里香种子萌发情况, 得出干旱与低温的交互作用显著影响了法国百里香种子的萌发。在不适宜的温度与干旱胁迫的交互作用下会使种子降低内含物, 进而影响其发芽率、延迟发芽时间, 甚至使种子完全丧失活力。浓度为30%的PEG-6000与8~10 ℃和12~14 ℃的交互情况下法国百里香种子无法萌发, 但是, 当温度上升为22~24 ℃的情况下, 在培养的第18天种子开始萌发, 最终发芽率仅为9%。说明低温不仅在一定程度上会引起种子的休眠, 而且随着温度下降, 种子的抗旱性也逐渐降低。
法国百里香种子的萌发与幼苗生长对温度和模拟干旱表现了不同的响应, 综合各指标表明当温度为12~14 ℃、PEG-6000浓度为10%的条件利于法国百里香种子的萌发, 而在幼苗生长过程中, 利于根、茎生长的适宜条件是22~24 ℃、无PEG-6000的处理。
The authors have declared that no competing interests exist.
| [1] |
|
| [2] |
|
| [3] |
|
| [4] |
|
| [5] |
|
| [6] |
|
| [7] |
|
| [8] |
|
| [9] |
|
| [10] |
|
| [11] |
|
| [12] |
|
| [13] |
|
| [14] |
|
| [15] |
|
| [16] |
|
| [17] |
|
| [18] |
|
| [19] |
|
| [20] |
|
| [21] |
|
| [22] |
|
| [23] |
|
| [24] |
|