




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
不同豆禾混播模式的草地生产性能
[祁军1 , 郑伟2, 3  , 张鲜花
, 张鲜花2, 3 , 唐高溶2 , 王祥2 , 朱进忠2, 3 ]
, 张鲜花|
|


第一作者:祁军(1978-),男,湖北房县人,助理研究员,本科,研究方向为作物栽培与育种。E-mail: [email protected]
选择5种豆科与禾本科牧草建植同行与异行豆禾混播草地,混播种类为2种豆禾牧草混播、5种豆禾牧草混播,豆禾混播比例为豆禾比6∶4、5∶5和4∶6。依据2012-2013年各混播组合的牧草产量、粗蛋白产量、粗脂肪产量、中性洗涤纤维产量、相对产量总和(RYT)及豆科牧草与禾草的相对密度和相对产量相异度,分析了不同混播群落空间结构下各混播处理的生产性能变化。结果表明,混2-1(鸭茅 Dactylisglomerata+红豆草 Onobrychisviciaefolia)、混2 -2(无芒雀麦 Bromusinermis+红豆草)的1∶1行处理具有较高的牧草产量与粗蛋白、粗脂肪和中性洗涤纤维产量,且均高于同行混播;同行混播牧草产量均低于异行混播。各混播处理RYT值均高于1,且混2-1、混2-2的1∶1行处理RYT值高于同行混播。2∶2行、3∶3行具有较高的相对密度和相对产量相异度,同行混播具有较低的相对密度和相对产量相异度。由此可见,从同行混播改为异行混播,可提高牧草产量、牧草品质和种间相容性,维持较高的群落稳定性,使豆禾混播草地的生产性能进一步增加。
The seven indexes including herbage yield, dissimilarity of grass and legume relative density, relative yield total (RYT), dissimilarity of grass and legume relative yield, crude protein yield, ether extract yield and neutral detergent fiber yield were selected to study the changes of the comprehensive production performance under different legume-grass mixtures in 2012―2013. The mixture patterns included two legume-grass combinations: 5, 2 species mixture at three sowing ratios of legume︰grass (6︰4, 5︰5, 4︰6), mixed cropping and intercropping respectively. The species for mixture include Onobrychisviciaefolia, Trifoliumpratense, Dactylisglomerata, Bromusinermis and Phleumpratense. The results showed that the forage yield, CP yield, EE yield, NDF yield and RYT values of legume-grass mixture (1∶1)were higher than mixed cropping in mixed 2 species-1 and mixed 2 species-1. The forage yield of mixed cropping were lower than intercropping in all species mixtures. The RYT values of all mixtures were higher than 1. The dissimilarity of grass and legume relative density, grass and legume relative yield for 2 rows legume︰2 rows grass and 3 rows legume︰3 rows grass were higher than 1 legume︰1 grass and mixed cropping. Therefore, from mixed cropping to intercropping like 1 legume︰1 grass, 2 rows legume︰2 rows grass and 3 rows legume︰3 rows grass, the forage yield and quality, the interspecific compatibility and the mixed community stability could improve. The comprehensive production performance of intercropping could further increase.
豆科与禾本科植物混播后对共同栖居的空间和各种资源进行竞争; 豆禾混播群落还能使土壤水分、养分和光资源的供给得到改善, 增加了这些资源的可利用性, 从而产生互利[1]。竞争和互利在豆禾混播群落中可同时存在, 两者的相对大小及对混播优势的贡献随间作/混播植物建植年限(作物的生育期)及空间结构变化而变化[2, 3]。在混播植物的整个生长季内, 当竞争作用强度大于互利时, 混播群落对资源的整体利用能力下降, 反之, 混播群落对资源的整体利用能力增加, 具有混播优势[4]。调控种间竞争与互利作用的相对大小对获得混播优势具有重要作用[4]。目前关于豆禾牧草混播优势的研究主要集中在物种组成、豆禾混播比例[5]、刈割强度[6]和施肥[7]等领域, 对群落空间结构的关注较少[3, 8]。群落空间结构可反映群落内各植物种的空间关系, 各植物种之间的竞争状况及空间生态位, 在很大程度上决定了群落组分的稳定性和发展的可能性[9]。与此同时, 许多学者将混播草地牧草产量等同于其生产性能, 将牧草产量的差异作为评价不同混播模式下草地生产性能优劣的最主要指标[10]。混播草地的生产性能应考虑草产量、牧草品质[11]、种间竞争关系[12]、群落稳定性[13]等诸多因素, 如果仅以牧草产量的差异来评价其生产性能, 进而判定混播优势的大小或劣势, 往往因割裂了各个因素对混播草地生产性能的综合影响而在一定程度上影响混播优势判定的科学性[14]。因此, 本研究以3种豆禾牧草种类组合和3种豆禾混播比例的同行混播与异行混播草地为研究对象, 以群落空间结构的差异为切入点, 对比分析其牧草产量、牧草品质、种间相容性和群落稳定性, 探索利用多指标综合评价系统评价豆禾混播草地的混播优势, 以期为利用空间结构调控来获得较高混播优势提供依据。
试验地点位于新疆维吾尔自治区伊犁哈萨克自治州昭苏盆地的昭苏马场山前草地(43° 07'33.12″N, 80° 59'56.18″E, 海拔1895m), 昭苏马场属温带山区半湿润易旱冷凉气候类型。年均温度2.7℃, ≥ 10℃年积温1416.8℃· d, 无霜期85~100d, 年均降水量512mm, 年均蒸发量1261.6mm。试验小区原为退化的山地草甸, 2011年重建成多年生豆禾混播草地。山地草甸植被主要有鸭茅(Dactylisglomerata)、无芒雀麦(Bromusinermis)、新疆鹅观草(Roegneriasinkiangensis)、绿草莓(Fragariavirdis)、草原糙苏(Phlomispratensis)、草原老鹳草(Geraniumpratense)、短柄苔草(Carexpediformis)等。盖度65%~95%, 草层高25~85cm, 牧草产量2500。土壤为黑钙土, 土壤有机质含量为10.32%~10.56%, 全氮、全磷和全钾含量分别为7.15、1.14和17.00g· kg-1, 碱解氮、有效磷和有效钾含量分别为1322.00、8.61和924.00mg· k。
试验设同行混播与异行混播两种混播方式(a因素, a=2), 混播组合设2种牧草混播(鸭茅+红豆草Onobrychisviciaefolia、鸭茅+无芒雀麦)和5种牧草混播(鸭茅+无芒雀麦+猫尾草Phleumpratense+红三叶Trifoliumpretense+红豆草)(b因素, b=3); c因素为混播比例(c=3), 豆禾比分别为6∶ 4、5∶ 5和4∶ 6; 5种牧草每种各设单播作为对照(CK=5); 3次重复, 随机区组设计, 共69个小区。每个小区面积均为3m× 4m, 行距15cm。鸭茅、红三叶单播的播量均为15kg· hm-2, 猫尾草单播的播量为12kg· hm-2, 无芒雀麦单播的播量为30kg· hm-2, 上述草种均为加拿大产; 红豆草单播的播量为60kg· hm-2, 品种甘肃红豆草(O.viciaefoliacv.Gansu), 为新疆本地产; 混播比例按种子占单播重量的实际用价来计算, 混播与单播密度相同, 播量及混播比例如表1所示。2011年5月15日播种, 建植当年花期(8月20日)刈割1次。试验期间不施肥、无灌溉, 每年春季人工除杂草2次。
| 表1 豆禾混播草地混播比例与播量 Table1 Mixed sowing ratio and sowing quantity in legume-grass mixture |
1.3.1 评价指标的选择 牧草产量:每年刈割1次, 均在牧草花期进行刈割(7月15日左右)。每个小区随机设置3个1m× 1m的样方(缺苗及边行不设样方), 样方内牧草按种留茬5cm刈割并称鲜重, 每种牧草取500g鲜样放入65℃恒温箱烘干称重, 计算干鲜比, 折算成干重。
牧草品质:在牧草产量测定的同时, 在每个小区按“ 十” 字法随机取500g牧草混合样, 3次重复, 混匀, 将烘干样品粉碎后, 过0.4mm筛, 测定牧草的粗蛋白(CrudeProtein, CP)、粗脂肪(EtherExtract, EE)、中性洗涤纤维(NeutralDetergentFiber, NDF)含量, 同时根据产草量, 换算成CP、EE和NDF产量[16]。
种间相容性:利用相对产量总和(RelativeYieldYotal, RYT)表述种间相容性[17], 因为RYT可从整体上说明混播植物间是否存在种间竞争[18], 且使用更为普遍[19]。
RYT= 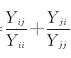
式中, Yij为种i与种j混播时种i的牧草产量; Yii为种i单播时的牧草产量; Yji为种j同种i混播时种j的牧草产量; Yjj为种j单播时的牧草产量[20, 21]。RYT< 1, 说明在混播组分种间竞争大于种内竞争; RYT=1, 说明混播组分种间和种内竞争相等; RYT> 1说明种间竞争小于种内竞争, 混播物种有某种程度的生态位分化, 其资源利用效率高于单播。
群落稳定性测定:草地群落稳定性指群落特征对于外界干扰因素的状态保持能力[14]。草地群落的稳定性主要表现在两方面, 一是群落结构的稳定性, 二是产量的稳定性。而群落某些特征的变化可以反映其稳定性[22]。
相对密度(Relativedensity, RD)可以理解为群落各组成结构对资源潜在的占有能力的大小, 相对产量(Relativeyield, RY)则是物种对已占有资源量的评价[23, 24, 25]。通过对两者“ 相异度(Dissimilarity)” (即干扰后与干扰前群落的相异程度)的评定, 可以定量反映出群落稳定性的变化[14]。
RDg=Dgl/pDg;
RDl=Dll/qDl;
RYg=Ygl/pYg;
RYl=Yll/qYl.
CD= 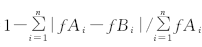
式中, RDg是禾本科相对密度, Dgl是混播下禾草分蘖数, Dg是单播下禾草的分蘖数, p是禾草的播种比例; RDl是豆科相对密度, Dll是混播下豆科牧草的分枝数, Dl是单播下豆科牧草的分枝数, q是豆科牧草的播种比例[23]。RYg是禾本科相对产量, Ygl是混播下禾草的产量, Yg是单播下禾草的产量; RYl是禾本科相对产量, Yll是混播下豆科牧草的产量, Yl是单播下豆科牧草的产量[26]。CD为相异度系数; fAi表示干扰前的群落特征值(其特征数为i=1, 2, 3, …, n, 这里取种植2011年的RD或RY); fBi为干扰后的群落特征值(这里取2013年的RD或RY)。当CD=1时, 说明群落有极强的稳定性; 而当CD≤ 0时, 说明其稳定性在这一外力作用下极差, 以至于完全丧失这一特征属性; CD值的大小可反映干扰力对群落的影响程度, 也就是草地的稳定性[12]。
1.3.2 评价模型的构建 应用模糊数学的隶属函数法对不同混播群落结构下豆禾混播草地的生产性能进行综合评价。首先对参评各因子值进行标准化处理:即每一因子数值除以该因子的最大值, 再乘以1000, 得到其标准化值[27]。然后利用模型U(Xik)=(Xik-Xkmin)/(Xkmax-Xkmin)计算牧草产量、牧草品质、种间相容性和群落稳定性4项指标的隶属度。最后采用上述4项指标隶属度的均值对豆禾混播草地的生产性能进行综合评价。其中, U(Xik)为第i种混播群落第k项指标的隶属度, Xik为评价指标的标准值或多项参评因子标准值的平均值; Xkmax、Xkmin分别为所有混播群落第k项指标最大值和最小值。
利用IBMSPSSStatistics21中的One-wayANOVA对不同混播群落结构的各指标进行方差分析、计算标准误, 并利用LSD对指标进行处理间比较, 差异显著性水平为0.05。
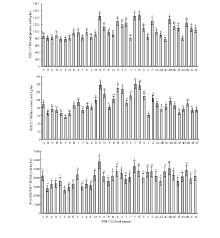
从混播组合(图1)来看, 组合N、AF牧草产量较高, 显著高于其他组合(P< 0.05); 组合B、C、E、F、G、J、L牧草产量较低, 显著低于组合R、S、V、X、AD、AH和AJ(P< 0.05)。从混播种类(图2)来看, 混2-1(红豆草+鸭茅)和混2-2(红豆草+无芒雀麦)牧草产量高于混5。从混播群落结构来看, 各混播种类的1∶ 1行牧草产量显著高于同行和3∶ 3行(P< 0.05); 混5和混2-1的2∶ 2行牧草产量低于1∶ 1行, 高于3∶ 3行, 但差异不显著(P> 0.05); 混2-2的2∶ 2行、3∶ 3行和同行混播牧草产量无显著差异(P> 0.05)。
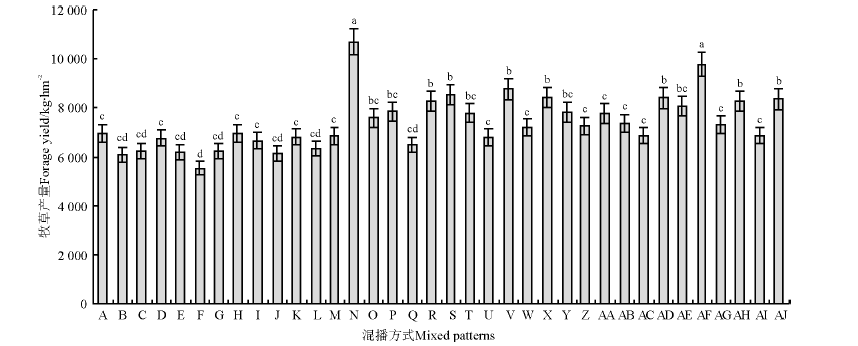 | 图1 不同混播组合牧草产量间的比较Fig.1 Forage yield of different cropping patterns and spatial structures 注:A, B, …, AJ分别代表混播比例和组合, 同表1所示。不同小写字母表示不同混播组合间差异显著(P< 0.05)。图3、图5、表2、表4同。 Note: A, B, …, AJ respectively represent the mixed combinations of mixed species and ratios, and were same table 1. Different lower case letters mean significant differences under different mixed combinations at 0.05 level. The same in Fig.3., Fig.5., Table 2, Table 4. |
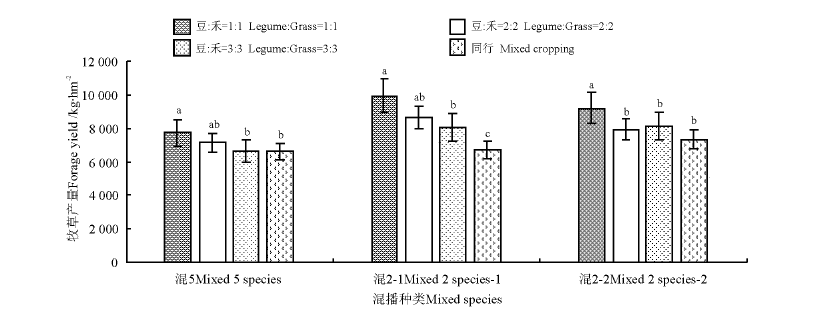 | 图2 不同混播空间结构牧草产量间的比较Fig.2 Forage yield of different mixed spatial structures |
注:不同小写字母表示同一混播种类不同混播空间结构间差异显著(P< 0.05)。下同。
Note: Different lower case letters for the same mixed species mean significant difference among different mixed spatial structures. The same below.
从混播组合(图3)来看, 组合N、V、W、Z和AD粗蛋白(CP)产量较高, 组合B、C、E、F、G、J、L、U、Y、AC和AG的CP产量较低。从混播种类(图4)来看, 混2-1和混2-2的CP产量高于混5。从混播群落结构来看, 混2-1和混2-2的1∶ 1行CP产量显著高于同行和3∶ 3行(P< 0.05); 混5的各处理间CP产量差异不显著(P> 0.05); 混2-1的2∶ 2行CP产量也显著高于3∶ 3行和同行(P< 0.05), 而混2-2的2∶ 2行和3∶ 3行CP产量则显著高于同行。从混播组合来看, 组合N、R、S、V和W粗脂肪(EE)产量较高, 组合F、Y的EE产量较低。从混播种类来看, 混2-1和混2-2的EE产量高于混5。从混播群落结构来看, 混5、混2-1和混2-2的各处理间EE产量差异不显著(P> 0.05); 混5的同行EE产量高于1∶ 1行、2∶ 2行和3∶ 3行(P> 0.05); 混2-1和混2-2处理EE产量则是1∶ 1行、2∶ 2行较高。从混播组合来看, 组合F中性洗涤纤维(NDF)产量最低, 组合N和V的NDF产量较高。从混播种类来看, 混5的NDF产量低于混2-1和混2-2。从混播群落结构来看, 混5同行的NDF产量显著高于其它混播群落结构(P< 0.05), 1∶ 1行、2∶ 2行和3∶ 3行之间则无显著差异(P> 0.05); 混2-1和混2-2的1∶ 1行NDF产量显著高于其它混播群落结构(P< 0.05), 同行、2∶ 2行和3∶ 3行之间则无显著差异(P> 0.05)。
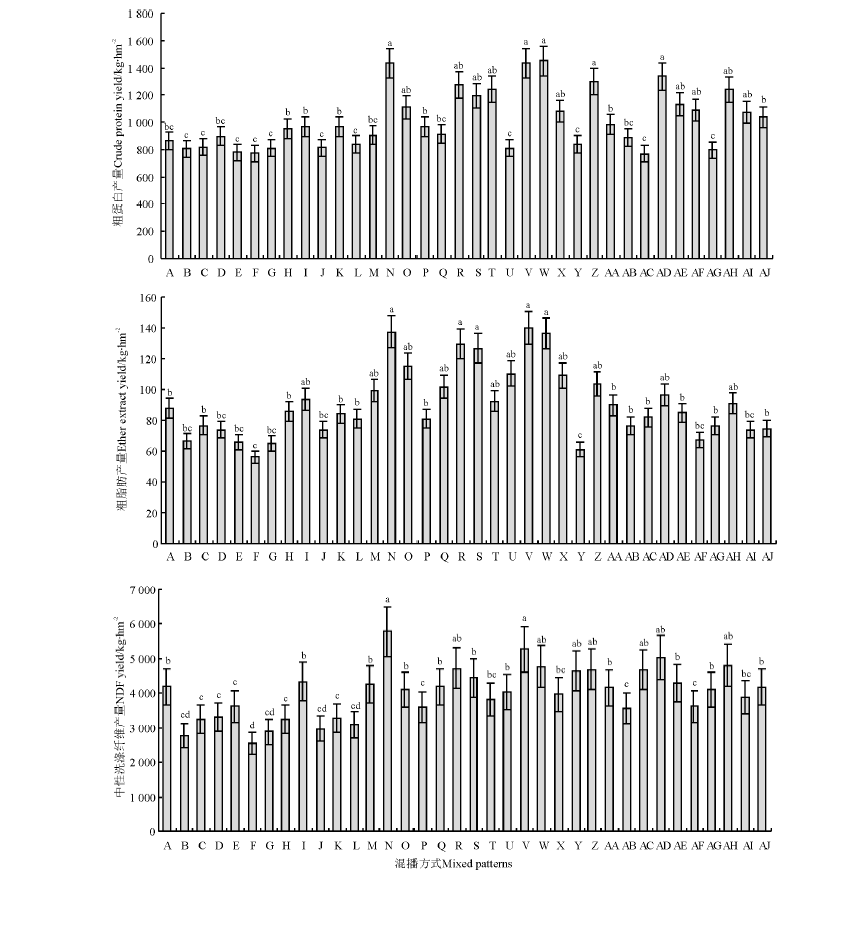 | 图3 不同混播组合牧草营养物质产量间的比较Fig.3 Forage nutrient yield of different cropping patterns |
 | 图4 不同混播空间结构下牧草营养物质产量间的比较Fig.4 Forage nutrient yield of different mixed spatial structures |
所有混播组合的相对产量总和(RYT)值均高于1。从混播组合(图5)来看, 组合N和AF的RYT值较高, 可达3.36以上, 与组合O、P、R、S、T、V、X、Y、Z、AB、AD、AE、AH和AJ无显著差异(P> 0.05), 但显著高于其它组合(P< 0.05)。从混播种类(图6)来看, 混2-1和混2-2的RYT值高于混5。从混播群落结构来看, 混2-1的1∶ 1行的RYT值显著高于同行(P< 0.05), 但2∶ 2行和3∶ 3行RYT值与其它混播群落结构间无显著差异(P> 0.05); 混5和混2-2各处理间的RYT值差异不显著(P> 0.05)。
 | 图5 不同混播组合相对产量总和的比较Fig.5 Relative yield total of different cropping patterns |
 | 图6 不同混播空间结构相对产量的比较Fig.6 Relative yield of ifferent mixed spatial structures |
从混播组合来看(表2), 组合T、AJ的豆科牧草相对密度相异度(CD-RDl)值较高, 组合Q、W、Z、AC的CD-RDl值较低。组合K、L、R、S、Z和AD的禾草相对密度相异度(CD-RDg)值较高, 组合Y、A、E和U的CD-RDg值较低, 与大部分组合差异显著(P< 0.05)。组合N、AF的豆科牧草相对产量相异度(CD-RYl)值较高, 组合U、B、K、W、AG的CD-RYl值较低, 与大部分组合差异显著(P< 0.05)。组合B、G、H、K、L、P、AB、AD、AE和AF的禾草相对产量相异(度CD-RYg)值较高, 组合A、E、I、M、Q、Y和AC的CD-RYg值较低, 显著低于其它组合(P< 0.05)。
| 表2 不同混播组相对密度和相对产量相异度的比较 Table2 Dissimilarity for relative density and yield of different cropping patterns |
从混播群落结构来看(表3), 混5同行的CD-RDl值显著高于1∶ 1行的(P< 0.05), 2∶ 2行、3∶ 3行的CD-RDl值则与其它混播结构无显著差异(P> 0.05); 2∶ 2行、3∶ 3行的CD-RYg值显著高于其它混播结构(P< 0.05), 同行的CD-RYg值则显著低于其它混播结构(P< 0.05)。混2-1和混2-2的3∶ 3行CD-RDl值显著高于其它混播结构(P< 0.05), 混2-1同行的CD-RDl值显著低于其它混播结构(P< 0.05), 而混2-2处理1∶ 1行的CD-RDl值最低; 混2-2同行的CD-RYg值显著低于其它混播结构(P< 0.05), 混2-1处理2∶ 2行CD-RYg值显著高于其它混播结构(P< 0.05), 而混2-2处理3∶ 3行CD-RYg值显著低于1∶ 1行(P< 0.05)。混5同行的CD-RYl值显著高于1∶ 1行和2∶ 2行(P< 0.05), 与3∶ 3行的无显著差异(P> 0.05); 同行的CD-RYG值显著低于其它混播结构(P< 0.05)。混2-1和混2-2处理3∶ 3行的CD-RYl值均显著高于2∶ 2行和同行(P< 0.05), 混2-1的1∶ 1行CD-RYl值则与3∶ 3行无显著差异(P> 0.05); 混2-1和混2-2同行的CD-RYg值显著低于其它混播结构(P< 0.05), 混2-1处理1∶ 1行的CD-RYg值显著低于2∶ 2行和3∶ 3行(P< 0.05)。从混播种类来看, 混2-1的CD-RDl值和CD-RYg值较高, 而混2-2的CD-RDl值较低, 混5的CD-RYg值较低; 混5的CD-RYl值低于混2-1和混2-2, 混2-2的CD-RYg值低于混2-1和混5的。
| 表3 不同混播空间结构下相对密度和相对产量相异度的比较 Table3 Dissimilarity for relative density and yield of different mixed spatial structures |
综合各个混播组合生产性能评价指标的标准值或多项参评因子标准值的平均值, 分别计算牧草产量、牧草品质、种间相容性和群落稳定性的隶属度, 以这4个隶属度的均值综合评价各混播组合的生产性能。组合N、AF的生产性能较高, 其隶属度在0.73以上; 组合A、C、Q、U、J、B、AC、E和F生产性能较低, 其隶属度在0.30以下(表4)。从混播种类来看, 混5的生产性能较低, 其隶属度值低于混2-1和混2-2。从混播群落结构来看, 同行的生产性能较低, 各个混播种类处理下其隶属度值均低于异行混播。
| 表4 豆禾混播草地综合生产性能指标的隶属度值及均值 Table4 Subordinate function values of four production performance indexes and their means of different mixed patterns |
混播植物的枝条、叶片、根系在空间上的分布差异或时间上的生育时期差异均可导致混播物种的生态位分离, 使混播牧草间的竞争强度降低, 实现光、水分和养分的利用互补, 提高草地生产力[28, 29]。同行混播时, 豆科牧草与禾草种植在一起, 地上部豆科牧草与禾草的叶片对光照形成激烈的竞争, 特别是禾草抽穗后, 其上部的叶片位置较高, 对下部的豆科牧草形成遮阴, 造成了豆科牧草光合作用强度下降[30]。而异行混播下, 豆科牧草与禾草的叶片有一定距离, 位置较高的禾草遮阴效果有限, 那么对光资源的竞争就没有同行混播那么激烈。本研究中, 同行混播牧草产量显著低于1∶ 1行(P< 0.05), 且低于其它异行混播处理。在混播种类和混播比例相同的情况下, 营养物质产量与牧草产量具有较高的相关性[12], 因此, 除混5外, 其它混播种类下, 也是同行混播营养物质产量低于异行混播。
与此同时, 豆禾牧草混播后, 豆科牧草与禾草根系的相互作用有效促进了系统对氮磷的吸收和利用[29, 31]。同行混播、1∶ 1行处理下豆科牧草的根系能够伸入到禾草根系区, 通过生物固氮和磷的高效利用减少了养分的竞争, 使得其表现出混播优势; 而3∶ 3行使禾草根系远离了豆科牧草, 地下养分资源无法高效利用, 使其牧草产量和营养物质产量均表现较差(图1、2)。由此可见, 1∶ 1行的异行混播有利于调控地上与地下资源的竞争, 使混播的豆禾牧草获得较大的牧草产量和营养物质产量优势。
对栽培草地而言, 增加群落物种数量或功能群数量往往能使栽培草地更有效地利用环境资源, 维持长期较高的生产力和稳定性[32]。本研究中, 各混播组合相对产量总和均高于1。这表明豆禾牧草混播时, 混播物种所采取的生态对策是其共用的资源在时间和空间上可协调分配, 种间竞争下降[28]。但混播物种间的互利提高群落生产力被认为在很低的物种丰富度水平上就可以达到饱和[33], 因此豆禾混播草地不可能无限地追求混播种类的多样性[34]。本研究中混5各处理豆科牧草与禾草相对密度、相对产量相异度均值均低于混2, 因此, 群落稳定性未随着混播种类数量的增加而上升。从混播组分来看, 豆科牧草相对密度相异度低于禾草; 除同行混播外, 豆科牧草相对产量相异度也低于禾草。由此可见, 豆科牧草的竞争力和耐刈割性均弱于禾草[35], 在混播群落中属于竞争力弱、持久性差的组分, 而禾草属于竞争力强, 持久性好的组分。从混播群落结构来看, 3∶ 3行豆科牧草与禾草相对产量及豆科牧草相对密度的相异度较高, 2∶ 2行禾草相对密度相异度较高, 同行混播禾草相对密度和相对产量相异度较低。因此, 种间竞争较弱的1∶ 1行、2∶ 2行、3∶ 3行具有较高的群落稳定性, 种间竞争较强的同行混播具有较低的群落稳定性。
从牧草产量、牧草品质、种间相容性和群落稳定性4个方面测度与比较了36个豆禾混播组合的生产性能。从牧草产量和品质来看, 混2-1(鸭茅+红豆草)、混2-2(无芒雀麦+红豆草)的1∶ 1行处理具有较高的牧草产量和牧草品质, 且均高于同行混播。从种间相容性来看, 各混播处理相对产量总和均高于1, 且混2-1、混2-2的1∶ 1行处理高于同行混播。从群落稳定性来看, 种间竞争较弱的1∶ 1行、2∶ 2行和3∶ 3行具有较高的群落稳定性, 种间竞争较强的同行混播具有较低的群落稳定性。由此可见, 从同行混播改为异行混播, 可提高牧草产量、牧草品质和种间相容性, 维持较高的群落稳定性, 使豆禾混播草地的生产性能进一步增加。
(责任编辑 王芳)
The authors have declared that no competing interests exist.
| [1] |
|
| [2] |
|
| [3] |
|
| [4] |
|
| [5] |
|
| [6] |
|
| [7] |
|
| [8] |
|
| [9] |
|
| [10] |
|
| [11] |
|
| [12] |
|
| [13] |
|
| [14] |
|
| [15] |
|
| [16] |
|
| [17] |
|
| [18] |
|
| [19] |
|
| [20] |
|
| [21] |
|
| [22] |
|
| [23] |
|
| [24] |
|
| [25] |
|
| [26] |
|
| [27] |
|
| [28] |
|
| [29] |
|
| [30] |
|
| [31] |
|
| [32] |
|
| [33] |
|
| [34] |
|
| [35] |
|