{kind=link}
大尤尔都斯高寒草原退化演替序列的划分
[张爱宁1 , 安沙舟1  , 荀其蕾
, 荀其蕾1 , 董磊2 , 董乙强1 , 苏清荷3 ]
, 荀其蕾|
|


第一作者:张爱宁(1989-),女,新疆和田人,在读博士生,主要从事草地资源与生态研究。E-mail:[email protected]
采用系统聚类分析法对巴音布鲁克大尤尔都斯紫花针茅( Stipa purpurea)高寒草原进行退化演替序列的划分,并对不同退化草地的植物群落结构进行比较。结果表明,随着草地退化演替程度的加剧,群落结构与主要植物种的优势地位发生明显变化,顶级植被紫花针茅的高度、盖度、密度及地上生物量均有所下降;退化演替模式为:紫花针茅+艹洽草属( Koeleria)→赖草属( Leymus) +委陵菜属( Potentilla)→苔草属( Carex) +马先蒿属( Pedicularis)→艹洽草属+委陵菜属;紫花针茅的重要值可作为划分巴音布鲁克大尤尔都斯高寒草地退化的重要指标;相同等级内各样地间相似性大,不同等级间样地相似性小。系统聚类的结果与各样地相似性比较的结果一致。
In the present study, the degraded succession series of Stipa purpurea in alpine grassland was classified using system clustering analysis in Yourdusi Bayanbulak, and grassland plant community structures with different degraded level were compared. With the increase of the grassland degradation succession degree, the plants community structure and the dominant position of the main plants obviously changed and the height, coverage, density and the above ground biomass of S. purpurea decreased. The degradation succession model of alpine grassland in Bayanbulak were S. purpurea+Koeleria→ Leymus+Potentilla→ Carex+Pedicularis→ Koeleria+Potentilla. The important value of S. purpurea was as an important index of grassland degradation in Yourdusi Bayanbulak. The similarity between samples from same level was higher while similarity between samples from different levels was lower. Clustering results consistent with various similarity comparison results.
紫花针茅(Stipa purpurea)草原是巴音布鲁克大尤尔都斯高寒草原上主要的草地类型[1]。它的分布范围极广, 不仅在维护自然生态系统稳定性方面起着重要作用, 同时还为畜牧业的发展提供了物质基础, 对巴音布鲁克高寒草原生态效益与社会经济效益具有重大意义[2]。但是由于不断增长的畜产品需求, 导致放牧超载严重, 草地退化情况非常普遍, 局部区域已经重度退化, 使得植物群落逆向演替, 造成草地生产力和总生物量的下降, 严重制约着当地草地畜牧业的发展[3]。因此, 对由于放牧造成的草地退化演替序列进行划分, 能够深入了解退化草地植被群落的结构与功能[4], 群落优势种的重要作用, 以及放牧干扰对草地退化演替的影响机制[5], 这在监测该地区的草地利用状况和恢复退化草地等方面具有一定的指导意义[4]。在草地退化演替序列的划分上, 前人已做了大量研究工作, 如以植物群落特征及植物多样性的变化来判定草地退化等级[6]; 基于净初级生产力的变化对草地退化进行监测研究[7]; 以植被生产力的降低作为草地退化演替的指标进行研究[8]; 以优势种羊草(Leymus chinensis)的数量作为聚类指标, 运用聚类分析对羊草草地退化演替草地进行划分[9]; 以生态因子指标作为聚类指标, 运用聚类分析对辽西低山丘陵区生态系统退化状况进行定量确定[10]; 建立植被与气候共同作用来判定草地退化等级的数据模型[11]。对草地退化的水分因素的研究表明, 土壤水分流失会导致草地退化[12]。划分退化草地等级大多采用定性指标, 也有少数学者采用定量指标进行划分[13], 草地植被的数量指标是评价草地退化程度的重要依据。巴音布鲁克大尤尔都斯高寒草地是当地主要的放牧草地。因此, 本研究以植被数量指标聚类分析的方法为主要切入点, 对巴音布鲁克大尤尔都斯高寒草地进行研究, 以期了解:1)巴音布鲁克大尤尔都斯高寒草地退化程度分为几个等级, 不同退化程度草地的演替模式与植被特征情况; 2)能否将紫花针茅的重要值作为划分此类型草地退化等级的重要指标?3)系统聚类的结果与各样地相似性比较的结果是否一致?
巴音布鲁克草原位于新疆巴州和静县西北, 伊犁谷底东南, 中部天山南麓, 东西长270 km, 南北宽136 km, 平均海拔约2 500 m, 四周山体海拔在3 000 m以上, 面积约2.3万km2, 是我国第一大亚高山高寒草甸草原[14]。巴音布鲁克草原由小尤尔都斯草原和大尤尔都斯草原组成[15], 草地总面积14 972.67 hm2, 可利用草地面积14 160.67 hm2, 其中大尤尔都斯草地面积9 430.67 hm2, 占总面积的63%, 小尤尔都斯草地面积5 540.67 hm2, 占总面积的37%[4]。巴音布鲁克地区年均气温-4.2 ℃, ≥ 0 ℃年积温1 341.5 ℃· d, ≥ 10 ℃年积温306.5 ℃· d, 极端最低气温-49.6 ℃, 年均风速2.8 m· s-1, 多年平均降水量280.5 mm, 年蒸发量1 132.4 mm, 全年积雪日数137 d, 最大积雪深度为45 cm, 属典型的高寒气候[16]。土壤类型为亚高山草原土[17]。紫花针茅+羊茅(Festuca ovina)草原是研究区域内典型的高寒草原类型[18], 分布范围在海拔2 460-2 760 m, 群落种类分属19科40属, 群落总盖度约50%[19]。
研究工作采取野外采样调查与室内统计分析相结合的方法进行。2015年8月在巴音布鲁克大尤尔都斯研究区(83° 10'-85° 50' E, 42° 40'-43° 10' N)进行实地取样, 采取完全随机设计。由于巴音布鲁克大尤尔都斯盆地中心区域为湿地, 草地类型为草甸草原, 不属于本研究的范围, 因此选择环绕湿地周边的典型紫花针茅高寒草原作为研究区域, 其原生植被主要是以紫花针茅为主。共取样地36个, 样地间距大于5 km, 坡向坡度基本一致, 最高海拔2 668 m, 最低海拔2 415 m, 平均海拔2 502 m。每个样地内取3个1 m× 1 m的样方, 样方间距大于2 m。记录样方内植物的物种名、高度、盖度、密度, 称量样方中不同植物的地上生物量。在研究区域内, 将国家级固定监测点XJW087(83° 45.216' E, 42° 50.544' N, 海拔2 444 m) 设为未退化的样地, 对其样地内的植物重要值进行排序, 由于其它5个属的植物只鉴定到属, 所以选择紫花针茅、羊茅属(Festuca)、艹洽草属(Koeleria)、委陵菜属(Potentilla)、苔草属(Carex)、赖草属(Leymus)6种植物的重要值为基础数据。
重要值IV=(Rh+Rc+Rd+Ry)/4, 其中Rh为草地植物相对高度, Rc为相对盖度, Rd为相对密度, Ry为生物学相对鲜重。
聚类分析(cluster analysis)是以分类对象的特征数量作为指标, 定量确定其分类对象之间的相似性, 并将相对同质的群组统计归为一类的统计方法[20]。因此以不同样地中6种植物的重要值作为数据, 采用SPSS 20.0软件, 选择系统聚类分析(hierarchical cluster) 模块, 聚类方法选择组之间的连接, 计算出各变量间的距离, 得到样本矩阵, 测量区间为平方Euclidean距离即每个变量之差的平方和, 最终得出树状聚类图。通过聚类图中的聚类距离(横轴数值)分析比较各数据间的相似程度, 从而将各样地归类, 根据归类结果结合植被特征指数划分草地退化等级。
各样地中紫花针茅重要值范围是0~0.51(表1), 其中14和20号样地中紫花针茅的重要值均为最大值, 其次是21号样地的紫花针茅重要值为0.50, 10、11、30、27号样地的紫花针茅重要值分别是0.43、0.41、0.38、0.34; 羊茅重要值范围是0~0.35, 其中6号样地的羊茅重要值最大, 其次是5、13、7号样地, 其羊茅重要值分别为0.33、0.3、0.29; 艹洽草重要值范围是0.02~0.25, 其中3号样地的艹洽草重要值最大, 其次为1、6号样地, 重要值分别为0.23、0.22; 委陵菜重要值范围是0~0.22, 其中9号样地最大, 为0.22, 其次是12、16号样地, 重要值分别为0.21、0.2; 苔草重要值范围是0~0.5, 其中17号样地最大, 其次是19、25、24号样地, 重要值分别为0.41、0.32、0.31; 赖草重要值范围是0~0.39, 其中12号样地最大, 其次是2、18、8号样地, 重要值分别为0.32、0.29、0.22。
| 表1 各样地紫花针茅、羊茅属、艹洽草属、委陵菜属、苔草属、赖草属的重要值 Table 1 The important value of Stipa, Festuca, Koeleria, Potentilla, Carex, Leymus in different sample plots |
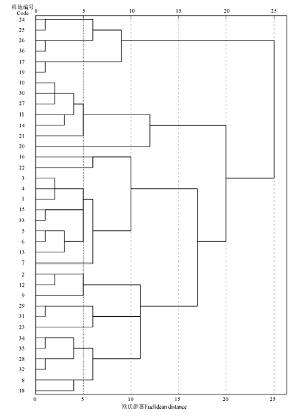
对36个样地进行聚类分析, 结果表明(图1), 在聚类距离为15时可将36个样地分为4类。由于27号样地是和静县国家级固定监测点XJW087(83° 45.216' E, 42° 50.544' N, 海拔2 444 m), 因此将监测点内植物群落设定为未退化群落。未退化草地为10、11、14、20、21、27、30号样地, 共7个样地, 地上生物量达到479.34 g· m-2, 盖度达到58.37%, 其中紫花针茅是优势种, 其地上生物量为55.69 g· m-2, 盖度为37.15%, 分别占群落的11.62%和66.71%。轻度退化草地为 2、8、9、12、18、23、28、29、31、32、34、35号样地, 共12个样地, 地上生物量为434.12 g· m-2, 植被盖度下降至53.86%, 其中紫花针茅生物量为25.85 g· m-2, 其盖度下降至15.65%, 分别占群落的5.96%和19.60%。中度退化草地为17、19、24、25、26、36号, 共6个样地, 由于长期受到家畜啃食、践踏等强烈干扰, 群落为适应恶劣环境改变了群落结构, 马先蒿和苔草取代紫花针茅成为优势种, 群落地上生物量和盖度比轻度退化略低, 但由于马先蒿等植物的入侵, 紫花针茅的正常生长受到抑制, 其地上生物量下降为20.67 g· m-2, 盖度下降为7.83%, 分别占群落的4.85%和14.57%。重度退化草地为1、3、4、5、6、7、13、15、16、22、33号样地, 共11个样地, 马先蒿、无芒雀麦(Bromus inermis)、细柄茅(Ptilagrostis)等植被的大量增加使紫花针茅的密度下降, 群落地上生物量降至220.11 g· m-2, 盖度降至45.39%, 紫花针茅的地上生物量和盖度也明显下降, 分别为8.00 g· m-2和5.77%, 占群落的3.63%和12.71%。
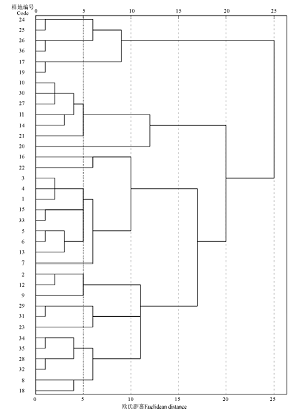 | 图1 各样地树状聚类图Fig.1 The cluster dendrogram of sample plots |
未退化草地各样地间的显著相似系数在0.84~0.97(表2)。轻度退化草地各样地间的显著相似系数为0.86~0.98, 与未退化群落之间的相似系数在-0.69~0.80。中度退化草地各样地之间的显著相似系数在0.83~0.99, 与轻度退化之间的相似系数在-0.90~0.06。重度退化草地各样地之间的相似系数在0.82~0.97, 与中度退化之间相似系数在-0.36~0.23。
| 表2 各样地相似性 Table 2 The similarity of sample plots |
巴音布鲁克大尤尔都斯紫花针茅高寒草地的退化演替模式为:紫花针茅+艹洽草→ 赖草+委陵菜→ 苔草+马先蒿→ 艹洽草+委陵菜, 随着退化演替程度的加剧, 群落主要物种的优势地位改变, 每个群落各草地主要植物种特征如表3所示。
| 表3 不同退化草地主要植物种类组成及特征 Table 3 The constitutes and characteristics of main plant species in different degraded stages |
在未退化阶段, 由于受放牧干扰较小, 紫花针茅+艹洽草为主的植物种类为群落优势种, 地上生物量较高, 群落垂直结构可分为3个层次:第1层以紫花针茅为主, 其平均高度达11.07 cm; 第2层以赖草、冰草、苔草、羊茅等占优势, 高度在5.85~8.47 cm; 第3层由低矮的艹洽草、委陵菜为主, 高度在2.97~3.43 cm。
在轻度退化阶段, 原先的群落优势种紫花针茅+艹洽草受到放牧干扰, 正常生长受到影响, 赖草、细柄茅成为优势种, 中层以紫花针茅和早熟禾为主, 虽然下层的艹洽草、委陵菜得到发展, 但毒害草马先蒿开始入侵群落, 垂直结构趋向简化。在中度退化阶段, 由于放牧过度, 家畜频繁啃食和践踏草地, 使得食口性较好的紫花针茅、羊茅等受到明显抑制, 而马先蒿、唐松草等毒害草的高度、盖度、密度均有所增加, 蒿属植物也开始入侵群落, 并且由于牲畜的频繁作用, 群落植被的高度、盖度、密度和生物量与轻度退化群落相比明显下降。
在重度退化阶段, 植被-土壤环境更趋恶化, 由于长期受到家畜的啃食与践踏作用, 重度退化阶段的植被较低矮, 植被稀疏, 生产力低。紫花针茅、羊茅等喜食性植物的发展更是受到严重制约, 而委陵菜、火绒草等植株矮小且适应干旱沙质环境的植物明显增多, 艹洽草、委陵菜成为群落优势种。
根据聚类分析的结果, 巴音布鲁克大尤尔都斯紫花针茅高寒草原退化演替过程可划分为4个等级, 即未退化草地、轻度退化草地、中度退化草地和重度退化草地。随着草地退化演替程度的加剧, 群落结构与主要植物种的优势地位发生明显变化, 顶级植被紫花针茅的高度、盖度、密度及地上生物量均有所下降。退化演替模式为:紫花针茅+艹洽草→ 赖草+委陵菜→ 苔草+马先蒿→ 艹洽草+委陵菜。退化演替过程中植被群落的垂直结构变化分上层、中层和下层。上层植被演替模式:紫花针茅→ 赖草+细柄茅→ 细柄茅→ 赖草; 中层植被演替模式:赖草+冰草→ 早熟禾+紫花针茅→ 马先蒿+无芒雀麦→ 早熟禾+紫花针茅; 下层植被演替模式:苔草+羊茅→ 艹洽草+马先蒿→ 苔草+紫花针茅→ 苔草+火绒草。紫花针茅的重要值可作为划分巴音布鲁克大尤尔都斯高寒草地退化的重要指标; 相同等级内各样地间相似性大, 不同等级间样地相似性小。系统聚类的结果与各样地相似性比较的结果一致。
草地退化的主要因素包括气候与人为, 本研究中过度放牧是导致草地退化的主要因素。随着牧压强度的变化, 草原植物群落的各植物种生态生物学特性及其功能地位发生变化[21]。随着草地退化程度加剧, 群落结构变化, 原生顶级植被紫花针茅的优势地位被最先入侵的赖草、细柄茅和早熟禾等禾本科草类所取代, 其原因可能是由于家畜通过采食行为和践踏作用导致土壤坚实[22, 23], 而赖草、细柄茅和早熟禾等小丛生禾草层以其强壮的根茎和更为广泛的生境适应性迅速繁衍, 成为独立的优势群落。小半灌木层片的蒿属植物在演替过程中以特异的气味来干扰动物的采食行为, 也增加其有性繁殖机会。随着牧压增大, 中层植被中逐渐出现了以毒素来保护自身不被动物采食的马先蒿、唐松草等毒害草[24], 可食性牧草种类减少[25]。重度退化时, 下层植被苔草、羊茅成为主要优势植被, 一方面由于植物高度下降限制了动物的采食行为, 另一方面丛生草层繁殖能力较强。紫花针茅原有优势地位完全被替代, 其生物量与盖度明显下降[26]。
在植物群落研究中重要值常被用于群落中物种数量分类及优势种集中趋势的分析[27]。前人研究多以植被特征数量值为划分草地退化等级的重要指标[28], 如冯秀等[29]以草地植物生物量作为评价草地退化等级的重要指标。而在本研究中发现, 以紫花针茅为优势种的植物群落, 36个样地中紫花针茅重要值排名前7位的与聚类结果中未退化草地完全一致, 轻度退化阶段有4个样地的紫花针茅重要值与划分结果一致, 中度与重度退化阶段各有两个样地的紫花针茅重要值与之相符。因此, 可将紫花针茅的重要值作为划分巴音布鲁克大尤尔都斯紫花针茅高寒草原退化等级的重要指标, 样地中其它主要植物可作为辅助划分依据。但是否适用于其它草地类型还有待验证。
根据系统聚类得出巴音布鲁克大尤尔都斯紫花针茅高寒草原可分为未退化草地、轻度退化草地、中度退化草地、重度退化草地4个退化等级。相同等级内各样地间相似性系数均呈正相关关系, 且相关系数平均在0.84~0.98, 而不同等级间的样地相似性系数相似性系数在-0.65~0.36。由此可得, 相同等级内各样地间相似性大, 不同等级间样地相似性小。这表明, 系统聚类的结果与各样地相似性比较的结果相一致。
研究中不足之处:一是以原生优势植物重要值作为划分草地退化等级的重要指标这一结论仅限于此次对巴音布鲁克大尤尔都斯紫花针茅高寒草原的研究中, 并未对其它相似类型草原作同样研究, 因此以原生优势植物重要值作为判定草地退化重要指标还有待其它研究结果的验证。二是未涉及土壤方面的研究, 无法评判土壤对草地退化方面的影响。
The authors have declared that no competing interests exist.
| [1] |
|
| [2] |
|
| [3] |
|
| [4] |
|
| [5] |
|
| [6] |
|
| [7] |
|
| [8] |
|
| [9] |
|
| [10] |
|
| [11] |
|
| [12] |
|
| [13] |
|
| [14] |
|
| [15] |
|
| [16] |
|
| [17] |
|
| [18] |
|
| [19] |
|
| [20] |
|
| [21] |
|
| [22] |
|
| [23] |
|
| [24] |
|
| [25] |
|
| [26] |
|
| [27] |
|
| [28] |
|
| [29] |
|