





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
俄罗斯木霉菌株GAU 1-X-2的生物学特性
[郭成1 , 张新瑞1  , 张军高
, 张军高2 , 李敏权1 , 徐生军1 , 王天玉2 ]
, 张军高, 徐生军|
|


第一作者:郭成(1985-),男,甘肃会宁人,助理研究员,在读博士生,主要从事玉米病害及微生物多样性研究。E-mail:[email protected]
从碳氮源、酸碱度及生长温度等方面对俄罗斯木霉(Trichoderma rossicum)菌株GAU 1-X-2的生物学特性进行了研究。结果表明,俄罗斯木霉菌株GAU 1-X-2的生长温度范围为10~35 ℃,最适温度为20 ℃;菌落在pH值为5.0~12.0的培养基上能够生长,pH为7.0时,生长最快,pH值为8.0时产孢量最大;营养生长最好的碳源为甘露醇,产孢量最好的碳源为葡萄糖;营养生长最适氮源为蛋白胨,在供试的7种氮源中,培养6 d未观测到该菌株产孢;完全黑暗条件有利于菌丝生长和产孢;孢子致死温度为68 ℃,10 min。
The biological characteristics of Trichoderma rossicum strain GAU 1-X-2 were examined using different carbon and nitrogen sources, pH, temperature, and other factors. The results indicated that T. rossicum could grow under temperatures of 10-35 ℃ and pH of 5.0-12.0, with optimal conditions of 20 ℃ and pH 7.0, although the highest sporulation occurred at pH 8.0. Mannitol was the optimum carbon source for hypha growth, but dextrose was the best for spore production. Peptone was the optimum nitrogen source for mycelia, but conidiophores were not observed after six days in the seven nitrogen sources. The best mycelia growth and the largest sporulation occurred with 24 h of darkness. The lethal spore temperature was 68 ℃ for 10 min.
随着化学农药的长期和大量使用而引起的农药残留、环境污染和病虫的抗药性等问题日益凸显, 给人民的生命安全和经济收入带来了巨大的风险。木霉属(Trichoderma )真菌作为土壤习居菌的主要类群, 其中有许多不同种和同一种不同菌株对植物病原真菌具有较强的抑制作用和拮抗效果, 目前, 已经应用于植物土传病害的生物防治[1, 2, 3]。木霉菌不仅能防治病虫害, 还能够增强植物抗逆性、提高营养利用效率、促进植物生长和修复农化污染环境等[4, 5]。因此, 利用木霉菌等微生物资源来防治植物病虫害已成为研究热点。
目前, 国内外学者不断搜集和筛选有较强抑制作物病原真菌的木霉菌株, 进行发酵培养和作用机制研究[6, 7, 8, 9, 10, 11, 12] , 包括竞争作用、重寄生作用、抗生作用、协同拮抗作用、诱导抗性及促进植物生长等。关于俄罗斯木霉(T. rossicum)的研究, 国外有文献报道其对核盘菌(Sclerotinia sclerotiorum)引起的大白菜(Brassica camperstris ssp. pekinens)菌核病有较好的防治效果[13]。国内崔岩等[14]从甘肃国营条山农场马铃薯(Solanum tuberosum)连作田植株根围土样中分离到1株木霉菌株GAU 1-X-2, 经形态学特征和ITS序列分析将其鉴定为俄罗斯木霉, 为国内木霉新记录种; 同时经过对峙培养法测定该菌株对马铃薯干腐病菌(Fusarium solani, F. sambucinum)和黑痣病菌(Rhizoctonia solani)具有较强的抑制作用。研究结果证明了俄罗斯木霉(T. rossicum)具有较好的生防应用潜力, 因此, 拟对俄罗斯木霉菌株GAU 1-X-2的生物学特性进行研究, 以期明确其营养生长及产孢所需的最佳碳源、氮源、pH、温度及光照条件等影响因子, 为下一步发酵培养俄罗斯木霉菌株GAU 1-X-2及其有效利用提供技术支撑。
俄罗斯木霉菌株GAU 1-X-2, 由甘肃农业大学草业学院植物病理研究室柴兆祥教授惠赠。
1.2.1 碳、氮源对俄罗斯木霉菌株GAU 1-X-2营养生长与产孢量的影响 将俄罗斯木霉菌株GAU 1-X-2在PSA平板上25 ℃下培养3 d, 用灭菌的内径6 mm的打孔器切取菌落边缘菌饼, 分别转移至以蔗糖、葡萄糖、果糖、麦芽糖、乳糖、可溶性淀粉、甘露醇制成的7种不同碳源的固体培养基中央[15, 16, 17, 18, 19, 20, 21]和以蛋白胨、牛肉膏、酵母膏、DL-天门冬酰胺、硝酸钾、尿素、硫酸铵制成的7种不同氮源的固体培养基中央[22], 25 ℃黑暗培养3 d。3 d后用十字交叉法测量菌落生长直径, 6 d后用血球计数板镜检孢子数量[23], 每处理 3 次重复。
1.2.2 温度对俄罗斯木霉菌株GAU 1-X-2营养生长与产孢量的影响 以直径6 mm的打孔器打取该菌株菌饼移至 PDA 培养基平板上, 分别置于 5、10、15、20、25、30和35 ℃共7个温度梯度下进行黑暗培养, 3 d后测量菌落生长直径, 6 d后镜检孢子数量, 每处理 3 次重复。
1.2.3 pH 值对俄罗斯木霉菌株GAU 1-X-2营养生长与产孢量的影响 利用0.1 mol· L-1的HCl和0.1 mol· L-1的NaOH将已灭菌的PDA培养基pH值分别调至为3.0、4.0、5.0、6.0、7.0、8.0、9.0、10.0、11.0和12.0共10个梯度, 依次制成不同pH值的PDA平板。移植直径6 mm的俄罗斯木霉菌株GAU 1-X-2菌饼于PDA平板中央, 25 ℃下黑暗培养, 3 d后测量菌落生长直径, 6 d后镜检孢子数量, 每处理3次重复。
1.2.4 光照对俄罗斯木霉菌株GAU 1-X-2营养生长与产孢量的影响 在智能人工气候箱(宁波海曙赛福实验仪器厂, PRX-450C)内设置12 h光暗交替(12 h光+12 h黑暗)、24 h连续光照(气候箱灯管为40 W日光灯)、24 h连续黑暗(无光照条件)共3种光照条件。25 ℃下培养, 3 d后测量菌落生长直径, 6 d后镜检孢子数量, 每处理3次重复。
1.2.5 俄罗斯木霉菌株GAU 1-X-2孢子的致死温度测定 第1步, 将2 mL孢子悬浮液分别置于40、45、50、55、60、65、70、75和80 ℃恒温水浴锅中处理10 min, 吸取20 μ L孢子悬浮液制成玻片, 置于25 ℃保湿黑暗培养, 每处理重复3次, 于12、24和72 h后镜检孢子萌发情况; 第2步, 根据第1步试验结果, 把2 mL悬浮液置于66、67、68、69 ℃恒温水浴锅中处理10 min, 同第1步观察孢子萌发情况[24], 同时吸取0.1 mL孢子悬浮液涂于PDA平板培养基上, 置于25 ℃培养箱内黑暗培养, 每处理重复3次, 5 d后检查PDA平板上菌落生长情况。
采用Excel 2010进行数据处理, 并采用DPS 2005软件进行单因素方差分析, Duncan氏新复极差法进行处理间差异显著性检验。
俄罗斯木霉菌株GAU 1-X-2在供试的7种碳源和7种氮源培养基上均能生长。生长最适的碳源为甘露醇, 平均菌落直径为75.3 mm, 显著大于其它6种碳源(P< 0.05); 其它6种碳源上的菌落直径为48.3~66.3 mm, 以淀粉和蔗糖为碳源培养基的菌落直径较低, 显著低于其它碳源培养基, 以葡糖糖、麦芽糖、乳糖和果糖为碳源的菌落直径差异不大(图1)。以蛋白胨、牛肉膏、酵母膏、天门冬酰胺为氮源, 菌落生长差异并不显著(图2), 以尿素为氮源培养基的菌落直径最低, 为5.0, 显著低于其它氮源培养基。
 | 图1 供试碳源对菌丝营养生长的影响Fig.1 Effects of different carbon sources values on mycelium growth注:不同小写字母表示处理间差异显著(P< 0.05)。下同。Note: Different lower case letters indicate significant difference at the 0.05 level. The same below. |
 | 图2 供试氮源对菌丝营养生长的影响Fig.2 Effects of different nitrogen sources values on mycelium growth |
GAU 1-X-2菌株适宜产孢的碳源为葡萄糖和麦芽糖, 1 mL孢子悬浮液平均产孢对数值为9.06~9.12, 其中最适碳源为葡萄糖; 其次为乳糖、果糖、蔗糖、甘露醇和可溶性淀粉的产孢量, 1 mL孢子悬浮液平均产孢对数值8.47~8.81, 葡萄糖和麦芽糖之间, 乳糖和果糖之间, 蔗糖、甘露醇和淀粉之间差异不显著(P> 0.05), 但它们之间产孢量差异显著(P< 0.05)(图3)。
 | 图3 碳源对产孢量的影响Fig.3 Effects of different carbon sources on sporulation quantity |
该菌株在10~35 ℃温度范围内均能生长, 平均菌落直径为1.3~71.2 mm。但在不同温度条件下, 该菌株菌落扩展速度不同, 适宜温度为20~25 ℃, 菌落平均直径为66.3~71.2 mm, 其中最适宜温度为20 ℃, 菌落平均直径高达71.2 mm。温度低于5 ℃, 菌落停止营养生长, 高于30 ℃菌丝生长速率减慢, 至35 ℃平均菌落直径为1.3 mm(图4)。在15~30 ℃的温度范围内该菌株均能产孢, 平均产孢量对数值为8.24~9.20, 适合产孢温度范围为20~30 ℃, 平均产孢量对数值为9.13~9.20, 其中最适宜产孢温度为20、25和30 ℃, 3个处理下产孢量差异不显著(P> 0.05), 但均显著高于15 ℃处理(P< 0.05)(图4)。
 | 图4 温度对菌丝营养生长及产孢量的影响Fig.4 Effects of temperature on mycelium growth and sporulation quantity |
pH为3.0时, PDA平板不凝固; pH为4.0时, PDA平板凝固较差。俄罗斯木霉菌株GAU 1-X-2在pH为5.0~12.0范围内菌丝均可进行营养生长, 对酸碱适应范围较广, 以pH为6.0~7.0菌丝营养生长较快, 其中最适pH为7.0, 平均菌落直径为66.3 mm; pH低于或高于7.0时, 菌丝生长速度开始下降(图5)。pH为8.0时的产孢量对数值为9.29, 显著高于除9.0以外的其它pH值(P< 0.05); pH为12.0时, 产孢量最低, 平均产孢量对数值为8.27(图5)。
 | 图5 pH对菌丝营养生长及产孢量的影响Fig.5 Effects of pH on mycelium growth and sporulation quantity |
俄罗斯木霉菌株GAU 1-X-2对光照条件要求不太严格, 在24 h黑暗、24 h光照、12 h黑暗+12 h光照条件下, 平均菌落直径分别为66.3、56.0和56.7 mm, 24 h光照和12 h黑暗+12 h光照处理间异不显著(P> 0.05), 但二者均菌落直径均显著小于24 h黑暗条件下的菌落直径(P< 0.05)(图6)。产孢量以24 h黑暗处理最高, 显著高于其它两个处理; 24 h光照最低, 显著低于其它两个处理(图6)。
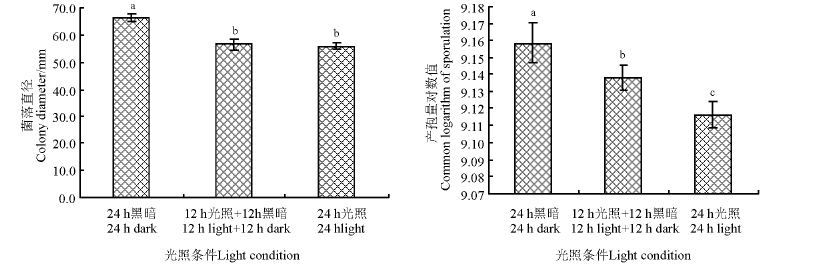 | 图6 光照对菌丝营养生长及产孢量的影响Fig.6 Effects of light on mycelium growth and sporulation quantity |
第1步试验结果显示, 俄罗斯木霉菌株GAU 1-X-2的孢子在40~65 ℃条件下处理10 min后仍能萌发, 而在70~80 ℃下不能萌发, 说明该菌株孢子的致死温度在66~69 ℃。第2步试验结果表明, 在66和67 ℃条件下, 孢子均能萌发, 5 d后能观察到有菌落生长, 菌落直径较小; 在68和69 ℃下未观察到有孢子萌发, 也未形成菌落, 说明该菌株孢子的致死温度是68 ℃, 10 min。
在木霉菌生物学特性方面, 许多学者做了大量研究[7, 22, 25, 26, 27, 28, 29], 而木霉菌的不同种及同种不同菌株在生物学特性方面存在差异。本研究结果表明, 俄罗斯木霉菌株GAU 1-X-2在以葡萄糖为碳源的培养基上产孢量最高, 与绿色木霉TR-8研究结果[26]一致, 但营养生长最适碳源为甘露醇, 与绿色木霉TR-8、哈茨木霉gz-2、棘孢木霉T31、长枝木霉TICC菌株营养生长最佳碳源分别为葡萄糖和麦芽糖、蔗糖、葡萄糖、果糖的研究结果[26, 27, 28, 29]有差异。有报道认为天门冬酰胺是绿色木霉TR-8菌丝生长和孢子产生的最佳氮源[26], 而本研究结果表明, 俄罗斯木霉菌株GAU 1-X-2营养生长最适氮源为蛋白胨, 在供试的7种氮源中, 培养6 d未观测到该菌株产孢, 究竟是何原因, 还有待于进一步试验。
俄罗斯木霉菌株GAU 1-X-2对温度和酸碱度的适应范围较宽, 其在10~35 ℃内均可生长, 菌丝最适宜扩展和产孢温度均为20 ℃; 菌丝最适营养生长的pH值为7.0, 产孢最适酸碱度为8.0, 与前人对绿色木霉、长枝木霉、棘孢木霉等的研究结果有差异[25, 26, 27, 28, 29], 哈茨木霉gz-2、棘孢木霉T31、绿色木霉TR-8菌株菌丝营养生长最适pH分别为5.0、6.0和7.0, 产孢最适pH依次为5.0、6.0和7.0, 而长枝木霉菌株TICC菌丝营养生长最适pH值为5.0, pH在3~7内均有利于产孢, 且各pH值间无显著差异。当pH 为3.0和4.0时, PDA平板不凝固或凝固较差, 未取得相关试验数据, 这并不代表该菌株在pH值低于5.0时不能生长, 需制成液体培养基进一步测试。本研究中, 俄罗斯木霉菌株孢子的致死温度为68 ℃、10 min, 表明该菌株具有一定的耐热性。这可能与本研究所用菌株分离来自景泰地区有关, 景泰县位于甘肃中部, 地处黄土高原与腾格里沙漠过渡地带, 特殊的生境决定了来自该地区的菌株在生物学特性等方面也存在特异性[27]。俄罗斯木霉菌株GAU 1-X-2在完全黑暗条件下, 菌丝营养生长最快, 产孢量最高, 该结果与光照处理对绿色木霉菌丝生长影响不明显的结果[26]较一致, 但可明显促进产孢的研究结果不一致, 这是否与菌株长期处于黑暗状态且适应其生存环境相一致, 需进一步开展相关研究。
本研究中的俄罗斯木霉菌株GAU 1-X-2为崔岩等[14]报道的国内新记录种, 本研究首次报道了其最佳碳源、氮源、温度、光照及酸碱度。但有关俄罗斯木霉菌株GAU 1-X-2对马铃薯干腐病菌和黑痣病菌的拮抗作用机理及发酵培养, 有待于进一步研究。
The authors have declared that no competing interests exist.
| [1] |
|
| [2] |
|
| [3] |
|
| [4] |
|
| [5] |
|
| [6] |
|
| [7] |
|
| [8] |
|
| [9] |
|
| [10] |
|
| [11] |
|
| [12] |
|
| [13] |
|
| [14] |
|
| [15] |
|
| [16] |
|
| [17] |
|
| [18] |
|
| [19] |
|
| [20] |
|
| [21] |
|
| [22] |
|
| [23] |
|
| [24] |
|
| [25] |
|
| [26] |
|
| [27] |
|
| [28] |
|
| [29] |
|