







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
一年生植物功能群对放牧草原生物量和土壤呼吸的影响
[梁茂伟1 , 梁存柱1  , 白雪
, 白雪1 , 苗百岭1, 2 , 王英舜3 , 包桂荣4 , 王譞1 ]
, 白雪|
|


第一作者:梁茂伟(1990-),男,内蒙古丰镇人,在读博士生,主要从事草地生态学和植被生态学研究。E-mail:[email protected]
作为草原群落重要层片的一年生植物功能群,在湿润季节和年份高度发育补偿群落生物量、维持放牧草原生态系统稳定,但尚不清楚一年生植物功能群通过这一补偿方式如何补偿群落地上/地下生物量和影响土壤呼吸。基于2012年和2013年内蒙古锡林郭勒大针茅(Stipa grandis)+羊草(Leymus chinensis)典型草原两个放牧强度试验样地群落地上/地下生物量、土壤呼吸及相关环境因子的测定。研究表明,1)一年生植物功能群对群落生产力的补偿作用及生长季土壤呼吸累积量在湿润年份(2012年)均大于干旱年份(2013年);且在中度放牧处理中均显著大于轻度放牧处理(P<0.05);2)土壤呼吸与一年生植物功能群地上、地下生物量、土壤5 cm温度和土壤0-10 cm含水率均具有极显著正相关关系(P<0.001)。由此表明,当放牧降低了草原群落多年生功能群生物量时,一年生植物功能群在湿润季节和年份可补偿群落生产力、促进土壤呼吸排放速率,进而影响草原生态系统碳循环过程。
, Bai XueAs a critical synusia in steppe grasslands, annual plant functional groups can compensate for changes in community biomass and maintain its stability in wet seasons and across years. This compensation should be evident because they are more sensitive to precipitation than perennial plant functional groups, especially in grazing communities with low biomass of perennial plant functional groups. However, it is not clear how the compensation effects of the annual plant functional groups improve productivity and affect soil respiration (Rs) in grazing areas in the steppe. We conducted a 2-year grazing experiment, including light-grazing (L) and moderate-grazing (M) treatments, in a typical steppe in Inner Mongolia in 2012 and 2013. Above-ground biomass (AGB), under-ground biomass (UGB), Rs, and environmental factors were monitored during the growing season. We found that: 1) either compensation of the annual plant functional groups occurred in the AGB and UGB, or Rs was higher in the wet year (2012) than in the dry year (2013); 2) both compensation effects in the annual plant functional groups in the AGB and UGB and Rs were significantly greater in moderate-grazing than light-grazing; 3) Rs was positively correlated with both the AGB and UGB of the annual plant functional groups, as well as with soil temperature and moisture (P<0.001). This study showed that grazing utilization decreased biomass of the perennial plant functional groups, and the annual plant functional groups dramatically developed in wet seasons and years to compensate for loss in community biomass to improve productivity and maintain ecosystem stability, which as a principal biotic mechanism that affects the ecosystem carbon cycle in grasslands.
碳循环是地球上主要的生物地球化学循环之一[1], 其中土壤呼吸是陆地生态系统碳循环中最重要的环节[2, 3]。天然草地作为全球分布面积最大的陆地生态系统, 80%的碳贮存在地下土壤库中[4], 因此土壤呼吸的动态将显著影响草地生态系统的碳储量[5]。
草原生态系统土壤呼吸主要是指土壤中产生CO2的所有代谢过程, 包括土壤微生物呼吸、活根系呼吸、土壤动物呼吸和含碳物质的化学氧化作用, 其季节动态主要受温度和水分及二者间配置的影响, 并且因群落类型和地理位置的不同而异[6]。其中植物根系呼吸和土壤微生物呼吸是草地生态系统土壤呼吸的主要组成部分。采用根系生物量梯度外推法在北美高草草原群落测得根系呼吸占土壤总呼吸的比例为40%[7]。采用除去根系法在澳大利亚一个多年生栽培草地测得根系呼吸量占土壤总呼吸量的53%[8]。Raich和Tufekcioglu[9]结合多数试验的测定结果, 并对各种因素加以考虑, 估计草原群落根系呼吸量占土壤总呼吸量的比例在17%~40%。采取根系生物量外推法在内蒙古典型草原测得根系呼吸占土壤总呼吸的15%~37%, 平均为24%[10], 并发现植物根系和土壤微生物呼吸速率及其季节变化主要受土壤温度和水分条件的控制[11]。在相同区域的研究结果还表明, 根系呼吸对土壤呼吸的贡献约为37%[12]; 对锡林河流域3个草原群落的研究发现, 根系呼吸占土壤呼吸的比例大多介于40%~50%[13]。可见, 草原群落地下根系对草原生态系统土壤呼吸的贡献十分突出。
近年来, 我国草地生态系统土壤呼吸研究十分活跃, 在土壤呼吸动态及影响因子等领域取得了诸多研究成果[14, 15, 16]。但是针对放牧对天然草地土壤呼吸的影响, 存在较多争议。在高山、高寒草甸的研究表明, 放牧促进土壤CO2排放[17, 18]; 类似的研究结果在内蒙古草甸草原[19, 20]、荒漠草原[21] 和科尔沁沙质草原[22] 均得到验证。但在内蒙古典型草原的研究结果却表明, 放牧对土壤呼吸速率影响不大[10], 可能因为放牧对草地土壤呼吸的影响主要与群落类型和放牧强度有关[23], 也可能是家畜采食、踩踏和排泄等活动间接影响群落生物和非生物因素, 进而引起草地生态系统土壤呼吸发生改变[24]。可见放牧对土壤呼吸的影响并不是简单的促进或者抑制, 其表现较为复杂。
在干旱与半干旱区的草原生态系统, 群落中往往存在恒有的一年生植物功能群或一年生植物层片[25], 特别在多雨的季节或年份高度发育[26]。通常放牧会导致草原群落建群种(多年生植物功能群)生物量的降低, 但由于放牧削弱了建群种层片的竞争优势, 又未抑制一年生植物层片的发育[27], 从而使一年生植物功能群充分利用相对“ 过剩” 的空间和养分资源而发育良好, 补偿群落地上与地下生物量[26, 27, 28, 29], 维持群落的稳定性[27, 30]。然而, 放牧群落中一年生植物功能群的补偿作用对草原生态系统碳循环的影响机制尚不是很明确。为此, 本研究以内蒙古典型草原大针茅(Stipa grandis)+羊草(Leymus chinensis)群落为研究对象, 分别在2012和2013年植物生长季, 监测两个放牧梯度群落土壤呼吸、地上和地下总生物量、多年生草本功能群生物量和一年生草本功能群生物量的动态, 探讨一年生植物功能群在放牧群落中补偿群落生物量、维持草原生态系统稳定性的作用机理, 及其对草原生态系统碳循环的影响机制。这一研究有助于深入认识一年生植物功能群在维持草原生态系统稳定性过程中的重要作用, 同时对草地管理具有一定的指导意义。
试验样地设于内蒙古锡林浩特市国家气候观象台生态监测样地(44° 08' N, 116° 19' E, 海拔1 129 m)。气候为典型的大陆性气候, 年平均降水量为300 mm, 主要集中在6 月-9月植物生长季阶段; 年平均气温3 ℃, 最高气温31.8 ℃, 最低气温-34.9 ℃, ≥ 10 ℃ 的年积温为2 700.5 ℃· d; 土壤类型为栗钙土[31, 32]。植物群落为大针茅(Stipa grandis)+羊草(Leymus chinensis)典型草原, 群落中一年生植物功能群主要为刺穗藜(Chenopodium aristatum)、猪毛菜(Salsda collina)、狗尾草(Setaria viridis)、虎尾草(Chloris virgata)和画眉草(Eragrostis pilosa)等, 但在不同的年份和季节物种组成及生物量变异很大。2012年5月-9月植物生长季降水量为434.9 mm, 年总降水量约520 mm, 显著高于多年平均值(300 mm), 划分为丰水年, 即多雨年份; 2013年5月-9月植物生长季降水量为234.7 mm, 年总降水量约为270 mm, 稍低于多年平均值(300 mm), 划分为少雨年份[33, 34]。
试验样地设在自然放牧区域内, 选择两个放牧强度群落, 由畜群点辐射状设置放牧样地, 通过观察并根据样地距离棚圈的远近[27, 35] 和群落当中建群种的地上生物量[36]划分为中度放牧利用和轻度放牧利用强度样地, 分别命名为“ 中度利用样地(moderate grazing, M)” 和“ 轻度利用样地(light grazing, L)” [19, 37]。两个样地地形等生境条件基本一致, 相距50~100 m, 连续两年均在同一区域测定。
1.3.1 土壤呼吸测定 采用LI-8100土壤CO2分析仪(LI-COR, Lincoln, NE, USA)连接PVC土壤呼吸监测环进行土壤呼吸的测定。每一年度第1次试验测定前, 将PVC土壤呼吸监测环(直径为10 cm; 高度为7 cm)钉入土壤当中, 地上部分预留2 cm, 每次监测前一天进行检查, 确保环内无植物, 若有则齐地面剪除地上部分, 直到当年试验结束取出, 分别设置5个重复。每个日动态监测都在当日06:00 到次日06:00之间进行24 h动态测定, 均监测12轮, 每轮时间间隔2 h。每次测定, 仪器自动测量土壤呼吸值, 单位为μ mol· (m2· s)-1。2012年度监测的时间分别为7月27-28日、8月5-6日和9月8-9日, 共3次; 2013年度监测的具体时间分别为5月12-13日、6月21-22日、7月16-17日、8月16-17日和9月16-17日, 共5次。
1.3.2 生物量测定 群落地上生物量(above ground biomass, AGB)采用收割法, 分种测定其高度、丛幅、株丛数和生物量; 样方面积为1 m× 1 m, 10个重复, 取样后当晚称取每个物种的鲜重, 之后置于65 ℃ 鼓风烘箱烘烤24 h, 烘干后称取其干重。群落地下生物量(under ground biomass, UGB)使用根钻分层取样, 直径为7 cm; 在地上生物量的样方取样后, 斜对角钻取两钻合二为一, 10次重复; 分层为地下0-5、5-10、10-20、20-30、30-40、40-50和50-70 cm。取样后将土柱放在0.2 mm网眼的纱网袋里, 用水冲洗干净, 置于65 ℃的烘箱中烘干称重, 地下生物量统计时不区分死根与活根。地上生物量和地下生物量观测日期为同一天, 均在土壤呼吸动态监测的前一天。一年生植物功能群地下生物量根据其根冠比计算获得, 一年生植物刺穗藜和猪毛菜平均值为0.5, 小禾草类平均值为1.5[38]。
1.3.3 环境因子测定 试验区域年平均降水、年平均气温数据来源于试验样地所在的锡林浩特国家气候观象台, 地下5 cm土壤温度(temperature of soil, TS 5 cm)使用LI-8100 8100土壤CO2分析仪自带的温度探头监测, 土壤10 cm含水率(water content of soil, WS 10 cm)使用TRIME-PICO64(Time domain Reflectometry with Intelligent MicroElements)土壤水分测定仪监测, 均设置5个重复。
数据分析与处理应用Excel 2013、SPSS 19.0和Sigmaplot 12.0完成。采用t检验、单因素方差分析(One-way ANOVA)和LSD最小显著差异分析法对数据进行分析。相关性分析用 Person相关。
2012年多雨年份两个放牧利用强度群落的地上生物量均显著高于2013年的(P< 0.05)(表1)。2012年度, 群落地上生物量的峰值均出现在8月份(图1), 中度利用群落各月份的总地上生物量显著高于轻度利用群落(表1), 但中度利用群落一年生植物功能群平均地上生物量为230.21 g· m-2, 占群落地上总生物量2/3多, 多年生草本功能群平均地上生物量仅为62.96 g· m-2(图2); 轻度利用群落一年生功能群地上生物量为46.16 g· m-2, 仅占群落总地上生物量的1/4左右, 大针茅+羊草平均地上生物量较高, 为141.76 g· m-2。2012年度一年生植物功能群物种主要由刺穗藜和猪毛菜的一年生植物组成。由此可见, 去除一年生植物功能群, 轻度利用群落各月份多年生草本地上生物量仍显著高于中度利用群落(P< 0.05)(图2)。
 | 图1 2012和2013年不同放牧强度下总群落地上生物量的生长季动态Fig.1 Seasonal growth dynamics of community above-ground biomass under different grazing intensities in 2012 and 2013注:图中不同小写字母表示同一日期不同利用类型间差异显著(P< 0.05)。下同。Note: Different lower case letters the same date indicate significant difference among different utilization types at the 0.05 level. The same below. |
 | 图2 2012和2013年不同放牧强度下多年生草本功能群地上生物量的生长季动态Fig.2 Seasonal growth dynamics of grass community above-ground biomass under different grazing intensities in 2012 and 2013 |
2013年度, 少雨年份, 中度和轻度利用群落的总地上生物量较低, 且不管是总地上生物量还是去除一年生植物功能群的多年生草本地上生物量, 轻度利用群落6月、8月和9月显著高于中度利用群落(P< 0.05)(图1、图2)。一年生植物功能群地上生物量在中度利用群落当中占群落总生物量的比例仅约1/8, 而在轻度利用群落当中几乎为0。2013年度一年生功能群主要物种由狗尾草、虎尾草和画眉草等小禾草组成。
2012年度, 中度利用群落的地下生物量(0-70 cm总值)显著高于轻度利用群落(P< 0.05)(表1)。
| 表1 2012和2013年不同放牧强度下群落地上、地下生物量及一年生植物功能群地上、地下生物量和土壤呼吸特征 Table 1 Characteristics of community above-ground and under-ground biomass, above-ground and under-ground biomass in terms of annual plants functional group, and soil respiration of community under different grazing intensities in 2012 and 2013 |
2013年轻度利用群落的地下生物量(0-70 cm总值)显著高于2012年, 年内植物生长季平均值与中度利用无显著差异(P> 0.05), 仅在6、8和9月表现出显著差异(P< 0.05), 地下生物量峰值出现在6月份(图3)。
 | 图3 2012和2013年不同放牧强度下群落地下(0-70 cm)生物量的生长季动态Fig.3 Seasonal growth dynamics of community under-ground biomass under different grazing intensities in 2012 and 2013 |
一年生植物功能群地下生物量2012年中度和轻度利用群落分别115.10 和23.08 g· m-2, 2013年分别为11.28和0.03 g· m-2(表1)。在两年中, 中度利用强度的一年生植物功能群地下生物量显著高于轻度利用群落(P< 0.05), 2012年多雨年份的显著高于2013年少雨年份。而且, 一年生植物功能群的地下生物量主要分布在地下0―10 cm, 并以活根系为主。
2.3.1 土壤呼吸日动态 中度利用群落的土壤呼吸大于轻度利用群落(P< 0.05), 且2012年多雨年份日动态相同时段内的值均大于2013年少雨年份, 年际间日动态波动趋势稍有差异。尽管两年均表现为单峰型曲线, 但2012年峰值均出现在13:00, 而2013年最大峰值均出现在09:00; 2012年和2013年的最小值均出现在次日05:00(图4)。
 | 图4 2012和2013年不同放牧强度下群落土壤呼吸的日动态Fig.4 Diurnal dynamics of soil respiration of community under different grazing intensities in 2012 and 2013 |
2.3.2 土壤呼吸生长季动态 两个利用强度群落土壤呼吸速率年际间相同月份内差异较大。2012年峰值均出现在7月份; 最小值均为9月份所测得值; 中度利用强度均显著大于轻度利用强度的土壤呼吸速率(P< 0.05)。2013年土壤呼吸值生长季动态基本为单峰曲线, 中度利用峰值出现在8月份, 轻度利用为7月份, 最小值均出现在9月份; 除7月份外, 中度利用强度均显著大于轻度利用强度的土壤呼吸速率(图5)。
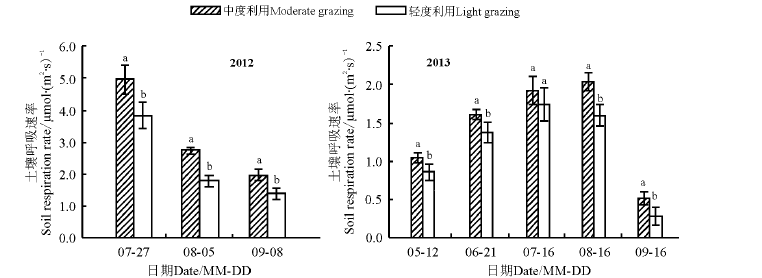 | 图5 2012和2013年不同放牧强度下总群落土壤呼吸的生长季动态Fig.5 Seasonal growth dynamics of community soil respiration under different grazing intensities in 2012 and 2013 |
放牧群落的土壤呼吸速率与一年生植物功能群地上、地下生物量的回归均是线性方程(地上, R2=0.203 6, P< 0.001; 地下, R2=0.162 9, P< 0.001), 即随一年生植物功能群地上、地下生物量的增大, 土壤呼吸速率极显著增强(图6)。
 | 图6 放牧群落土壤呼吸与一年生植物功能群AGB和BGB的线性回归Fig.6 Linear regression between soil respiration of grazing community and AGB and BGB for annual plants functional group |
放牧群落的土壤呼吸速率与一年生植物功能群地上生物量占总地上生物量的比例(R2=0.212 0, P< 0.001)和地下生物量占0-10 cm地下生物量的比例(R2=0.067 8, P< 0.05)的回归均是线性方程, 即随一年生植物功能群地上、地下生物量在群落中所占比例的增大, 土壤呼吸速率显著增强(图7)。
 | 图7 放牧群落土壤呼吸与一年生植物功能群ABG 占总AGB的比例和UGB占0―10 cm UGB比例的线性回归Fig.7 Linear regression between soil respiration of grazing community and annual plant functional group and UGB of total AGB and 0-10 cm UGB, respectively for annual plants functional group |
2012年和2013年的植物生长季, 两个群落的土壤呼吸与地下5 cm土壤温度和地下10 cm土壤含水率存在极显著的正相关性(P< 0.01)。尽管草地受放牧影响, 但不同利用强度的草地生态系统的土壤呼吸依然受温、湿度因子的控制, 表现为随着温度的升高和土壤湿度的增加, 土壤呼吸强度显著增加(图8)。
 | 图8 放牧群落土壤呼吸与土壤温度和土壤含水率的线性回归Fig.8 Linear regression between soil respiration of grazing community and soil temperature and soil water content |
在内蒙古典型草原区, 通常认为重度退化草地为冷蒿(Artemisia frigida)、糙隐子草(Cleistogenes squarrosa)群落, 中度退化草地为冰草(Agropyron cristatum)等群落, 轻度或未退化群落为羊草或大针茅群落, 群落的优势种均发生了更替[28, 29, 36], 本研究试验样地的群落优势种均未发生变化, 仅多生年优势物种生物量下降, 可能是由于短期高强度的放牧所导致, 所以本研究的中度放牧和轻度放牧仅代表两个不同放牧利用强度群落, 并未表现出典型的群落退化特征[29, 36]。
典型草原一年生植物功能群是适应于季节降水的不稳定层片, 由于典型草原退化群落多年生草本植物盖度和生物量均较低, 群落有相对“ 过剩” 的空间与养分资源, 在多雨的季节或年份, 一年生植物功能群发育良好, 而正常群落, 由于资源已被多年生草本群落较充分的利用, 即使降水较多, 一年生植物功能群也不会很发达[26, 27, 28]。因此, 在2012年降水量较大的情况下, 一年生植物在群落组成中占据很大一部分, 在中度利用群落中占78.52%、在轻度利用群落中占24.56%, 因而表现为中度利用的群落总地上生物量反而高于轻度利用群落。2013年降水较少, 接近正常年份, 群落一年生植物功能群不发育, 群落总生物量基本反映了放牧利用梯度的特点。包括一年生植物功能群在内的草原生态系统不同功能群之间具有相互补偿效应, 是导致草原群落具有相对稳定性的重要机制[30]。
一年生植物功能群多为“ 短命型” 的浅根系植物, 集中分布于0-10 cm浅层土壤, 而且多为活根系[25, 38]。基于典型草原群落地下生物量的动态监测发现, 在不区分活、死根系的情况下, 群落地下生物量(0-70 cm)约为2 344.72 g·
草地生态系统土壤根系呼吸占土壤总呼吸的40%~50%[7, 8, 9], 在典型草原的研究中也发现放牧群落的土壤呼吸受群落地下生物量的显著影响[10, 12, 13]。本研究中, 2012年和2013年中度利用群落的总地下生物量(0-70 cm)均大于轻度利用群落, 因此土壤呼吸速率也均高于轻度利用。其中, 放牧群落的土壤呼吸值与一年生植物功能群地下根量存在显著的正相关关系(图7)。野外实测的草地群落土壤呼吸主要受0-10 cm活根系的影响, 相对于轻度利用, 中度利用的一年生植物功能群较发达, 尤其是在多雨的2012年, 一年生植物功能群在补偿地上生物量的同时, 对地下生物量起到较大贡献, 主要表现在对地下0-10 cm活根系生物量的补偿, 而群落土壤呼吸当中的根系呼吸主要来源于植物活根系的呼吸, 因此, 一年生植物功能群通过对群落地下活根系生物量的补偿, 影响群落根系呼吸, 进而影响群落土壤呼吸。多雨年份, 一年生植物功能群不仅在典型草原的退化群落较发达, 在荒漠草原和草原化荒漠群落则更为发达[25]。由于忽略了一年生植物功能群的作用, 往往导致退化群落土壤呼吸表现出与放牧梯度的不一致性, 有时土壤呼吸速率与放牧强度表现为负相关关系[19, 20, 21], 有时影响不大[10], 也有研究发现放牧促进土壤CO2的排放[17, 18]。而本研究中则在多雨年份呈明显的正相关。另外, 土壤微生物的种类和数量与植物根系的空间分布和数量紧密相关, 也可能是影响群落土壤呼吸的重要生物因子之一[42]。当然, 放牧对草地土壤呼吸的影响较为复杂[24], 群落类型和放牧强度对其也有很大影响[23], 但不论放牧强度与群落类型如何, 群落的地下生物量是影响群落土壤呼吸的重要因子。
土壤水分也是影响微生物活性的重要环境因子, 土壤水分主要通过对植物和微生物的生理活动、微生物的能量供应与分配等调节和控制气体的释放, 实现对土壤呼吸影响的间接作用[43, 44]。草地生态系统土壤呼吸近70%受控于土壤温湿度, 土壤呼吸随温度升高而增加[11]; 当水分因子成为胁迫因子时, 与土壤含水率也有关系[45, 46]。本研究也发现两个利用类型的土壤呼吸速率对温湿度因子较为敏感, 存在极显著正相关关系, 与其他学者在典型草原上的研究结果基本一致[10, 11, 12, 13, 14, 15, 16, 17, 18, 19, 20, 21, 22, 23, 24, 25, 26, 27, 28, 29, 30, 31, 32, 33, 34, 35, 36, 37, 38, 39, 40, 41, 42, 43, 44, 45]; 另外, 土壤呼吸年际间生长季趋势的差异可能主要是年际间水热因子分布的不均衡所导致的[46], 这也可能是本研究中土壤呼吸年际间波动趋势存在差异的原因之一。可见, 即使在放牧利用过程, 温、湿度因子依然对草地生态系统碳循环有一定的调控作用。
总之, 本研究定位研究内蒙古典型草原大针茅+羊草群落, 通过揭示放牧群落中一年生植物功能群对地上、地下生物量的补偿以及对土壤呼吸的影响, 得出:放牧群落的多年生草本植物地上生物量随放牧强度的增加而减少, 但是由于一年生植物功能群在多雨的季节和年份对群落生物量的贡献较大, 群落总生物量表现出与放牧梯度不一致的特征, 从而维持群落稳定性; 而且, 放牧草原的土壤呼吸与一年生植物功能群的地上、地下生物量和土壤温、湿度环境因子存在显著的正相关关系, 当一年生植物功能群在多雨的季节和年份表现出较强的补偿效应时, 致使放牧群落的土壤呼吸随放牧强度的增加而增加。这一研究清晰揭示了一年生植物层片在草原生态系统中的生态功能, 阐明了放牧利用对草原生态系统碳循环的影响机制, 同时为退化草原合理利用与保护提供了一定的科学依据。
The authors have declared that no competing interests exist.
| [1] |
|
| [2] |
|
| [3] |
|
| [4] |
|
| [5] |
|
| [6] |
|
| [7] |
|
| [8] |
|
| [9] |
|
| [10] |
|
| [11] |
|
| [12] |
|
| [13] |
|
| [14] |
|
| [15] |
|
| [16] |
|
| [17] |
|
| [18] |
|
| [19] |
|
| [20] |
|
| [21] |
|
| [22] |
|
| [23] |
|
| [24] |
|
| [25] |
|
| [26] |
|
| [27] |
|
| [28] |
|
| [29] |
|
| [30] |
|
| [31] |
|
| [32] |
|
| [33] |
|
| [34] |
|
| [35] |
|
| [36] |
|
| [37] |
|
| [38] |
|
| [39] |
|
| [40] |
|
| [42] |
|
| [43] |
|
| [44] |
|
| [45] |
|
| [46] |
|