

{kind=link}
{kind=link}
{kind=link}
{kind=link}
呼伦贝尔典型草原的利用方式对地上啮齿动物群落多样性的影响
[白萨如拉1, 2  , 袁帅
, 袁帅1, 2 , 满都呼1, 2 , 武晓东1, 2 , 付和平1, 2 , 高全荣3 ]
, 袁帅, 付和平|
|


第一作者:白萨如拉(1988-),女(蒙古族),内蒙古锡林郭勒人,在读硕士生,主要从事草原护与环境方向研究。E-mail:[email protected]
2012年7、9月及2013-2014年5、7、9月,采用标志重捕法研究了连续放牧、季节轮牧和禁牧对呼伦贝尔典型草原啮齿动物捕获率、捕获量比例、多样性指数(丰富度指数、均匀性指数、Shannon-Wiener指数)的影响。结果表明,1)啮齿动物捕获率表现为连续放牧>季节轮牧>禁牧,连续放牧区显著高于禁牧区( P<0.05)。2)不同放牧方式使啮齿动物群落结构发生了一定的变化。连续放牧区主要以草原黄鼠( Citellus dauricus)为主,捕获量比例为38.60%;季节轮牧区以黑线仓鼠( Cricetulus barabensis)和草原黄鼠占的比例较大,捕获量比例分别为37.30%和31.50%;禁牧区以黑线仓鼠为主,捕获量比例为38.89%。3)连续放牧区啮齿动物丰富度指数、Shannon-Wiener多样性指数均为最高;季节轮牧中均匀性指数为最高;禁牧中丰富度指数、Shannon-Wiener指数、均匀性指数均为最低。连续放牧中的丰富度指数和Shannon-Wiener指数显著高于禁牧( P<0.05)。不同放牧方式明显改变了啮齿动物群落组成及多样性。综上,本研究结果表明,季节轮牧方式更利于合理利用草地资源并且能够抑制鼠害发生。
Field experiment with three grazing treatments, including continuous grazing, rotational grazing and forbidden grazing, was conducted to determine the effect of utilization pattern on rodent capture rate and its community diversity in the typical steppe of Hulun Buir from 2012 to 2014 by using live trap method. This study showed that capture rate of rodents was significantly higher in continuous grazing area than that in forbidden grazing area, and the grazing treatments altered the community structure of rodents, indicating that Citellus dauricus with 38.60% capture rate was dominant in continuous grazing area, and C. barabensis, and C. dauricus were dominant species in rotational grazing area, accounting for the 37.30% and 31.50% capture rate, respectively, and C. barabensis was dominant species in forbidden grazing area, with capture rate of 38.89%. This study also showed that the highest species richness and Shannon-Wiener index of rodent community were observed in continuous grazing area, and the highest evenness index of rodent community was found in rotational grazing area, while the lowest richness index, Shannon-Wiener index and evenness index of rodent community were observed in forbidden grazing area. The richness and Shannon-Wiener index in continuous grazing area were significantly higher than those in forbidden grazing area ( P<0.05). These results indicated that the rotational grazing pattern encouraged grassland resources to effectively utilize and controlled the rodent troubles.
近年来, 草地的不合理利用加快了草地生产力的退化[1]。放牧制度的不合理导致草地退化, 对于一些鼠类创造出适宜的生存环境, 而鼠类数量的增加又加重了草地的承载力, 使草地进一步的退化, 形成一种恶性循环[2], 从而破坏了草地生态系统原有的稳定性。作为草地上主要野生消费者的啮齿动物, 他们对环境的变化非常敏感, 栖息地环境的改变也必然导致啮齿动物群落发生变化[3, 4]。啮齿动物因分布广、对微环境变化反应敏感以及取样相对简易等特点一直以来都是生态学者们的研究对象[5, 6]。其群落多样性指数随环境的变化而发生改变, 因此可以通过群落多样性来反映栖息地的变化和人类活动干扰的程度[7]。
家畜的采食使草地生产力、植物群落结构、功能发生变化从而间接改变啮齿动物栖息地环境和食物资源[8, 9, 10]。家畜的践踏直接破坏啮齿动物的洞穴、增加土壤紧实度、降低土壤水势、影响啮齿动物地下洞穴的微气候环境、改变土壤理化性质。这种外部环境条件的变化对不同的啮齿动物种群产生相异的影响。对一些种类提供适宜的生境, 增加其种群数量, 而对于另一些种群有负面的影响, 阻碍其种群的发展, 从而改变啮齿动物群落组成、结构及群落多样性[11]。
近年来, 国内外对啮齿动物群落多样性的研究已有较多报道[12, 13]。但主要是对不同生境类型的啮齿动物群落多样性和多样性空间尺度进行了分析, 对于典型草原不同放牧方式对啮齿动物群落多样性的研究较少。本研究以围栏放牧模拟不同放牧方式, 通过测定啮齿动物捕获率、捕获量比例以及群落多样性指数来研究啮齿动物群落在不同放牧方式下的响应, 为探寻既能合理利用草地资源, 又能控制鼠害的放牧方式提供生态学依据。
研究区位于内蒙古呼伦贝尔新巴尔虎左旗新宝力格苏木巴彦希勒嘎查, 地理位置117° 33'-120° 12' E、46° 10'-49° 47' N, 草地类型为典型草原, 属中温带大陆性季风气候, 海拔628~672 m, 年平均气温-2~-0.5 ℃, 无霜期100~127 d, 平均年降水量为271.8 mm, 主要集中在7-8月, 主要植被有蒙古韭(Allium mongolicum)、黄蒿(Artemisia scoparia)、冷蒿(Artemisia frigida)、冰草(Agropyron cristatum)、糙隐子草(Cleistogenes squarrosa)、克氏针茅(Stipa krylovii)、羊草(Leymus chinensis), 有少量的艹洽草(Koeleria cristata)、麻黄(Ephedra monosperma), 土壤为栗钙土。
在试验样地中选择基底条件相似的草地通过围栏围封3个等面积(10 hm2)的样地, 分别设置连续放牧区(Continuing grazing)、季节轮牧区(Rotational grazing)和禁牧区(Forbidden grazing), 各样区间的间隔为800~1 000 m。本研究开始时间为2012年7月, 从7月到9月进行放牧。2013年开始在每年的5月到9月进行放牧试验。连续放牧为围栏内进行自由放牧, 季节轮牧样地分为两个等面积小样区(面积10 hm2), 在两个轮牧小样区间进行轮流放牧, 轮牧周期为两个月。3个样地动物数据的采集都在10 hm2面积中取样。禁牧区作为对照区不进行放牧(2012年7月开始禁牧)。连续放牧与季节轮牧在每10 hm2样地中放两头牛, 放牧试验动物皆为同龄3岁体重相等成年公牛, 每年5、7、9月初连续4 d进行动物数据的取样。
2012年7、9月和2013-2014年5、7、9月各处理区随机设置3个同心圆, 各同心圆布放半径分别为7、14、21 m, 每个同心圆布放25个活捕笼(图1), 活捕笼规格为27.5 cm× 12.5 cm× 12.5 cm, 采用标志重捕法捕获啮齿动物, 新鲜的花生米作为诱饵, 每次连续调查4 d, 每天上、下午各查笼1次, 记录所捕个体的种名、性别、体重、繁殖状况及捕获位置。使用标志重捕法收集啮齿动物数据, 标志方法为皮下注射PIT芯片。
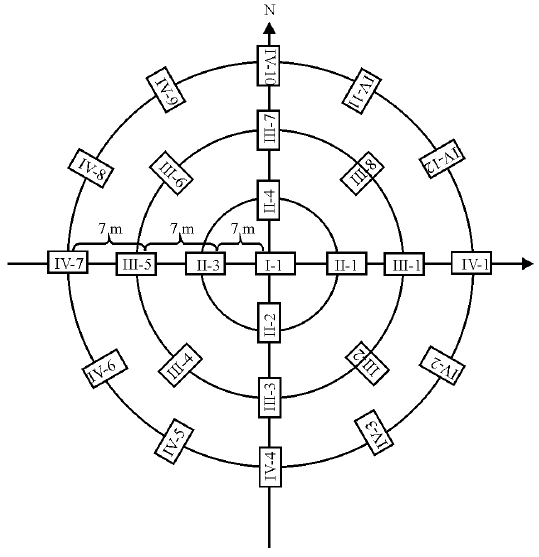 | 图1 圆型布笼方位图Fig.1 Location map of round cage |
2012年7月在试验开始前对各处理区植被情况(每个同心圆随机选取5个0.5 m× 0.5 m的植物样方, 并记录植物名称及植被高度、盖度、密度、生物量、枯落物干重)及啮齿动物数量进行了详细调查, 并通过方差分析(方差分析前对数据进行了正态分布检验, 均符合正态分布)比较不同样区间的植物及动物数据, 结果显示:各样区间的植被高度、盖度、密度、生物量及枯落物干重均无差异(P> 0.05)(表1)。啮齿动物总捕获率以及各个鼠种的捕获率在3种不同放牧方式下均无差异(P> 0.05)(表2)。
| 表1 基底植被调查(面积 0.25 m2) Table 1 Background survey of vegetation investigation(area 0.25 m2) |
| 表2 啮齿动物群落基底调查 Table 2 Background survey of rodent community |
对不同放牧方式下啮齿动物捕获率、捕获量比例、啮齿动物群落多样性指数采用以下方法测度:
(1)捕获率(百笼捕获率):以100笼所捕获的不重复啮齿动物计算。
捕获率=
(2)捕获量比例(pi):第i个物种的捕获数量与其所在群落中动物总捕获数量的比值。
(3)丰富度指数(Richness index), 计算公式为:
R=(S-1)/lnN。
式中, R为群落物种丰富度指数, S为物种数, N为群落中所有物种个体数。
(4)群落多样性指数(Diversity index), 采用Shannon-Wiener指数, 计算公式为:
H=-
式中, H为多样性指数, Pi为i个体在群落中的比例。
(5)均匀性指数(Evenness index)计算公式为:
J=H/Hmax, Hmax=lnS .
式中, J为均匀性指数, H为多样性指数, S为物种数。
通过SAS 9.0软件采用单因素方差分析(One-way ANOVA)对不同放牧方式下啮齿动物群落参数进行了差异性分析。(方差分析前对数据进行了正态分布检验, 均符合正态分布), 显著水平为0.05。应用Excel 2003软件完成数据的整理及构图。
2012年7、9月和2013-2014年5、7、9月在野外共布放了7 200个笼日, 捕获啮齿动物分属3科6属6种:1)松鼠科(Sciuridae), 草原黄鼠; 2)仓鼠科(Cricetidae), 黑线毛足鼠、黑线仓鼠、长爪沙鼠、布氏田鼠。3)鼠兔科(Ochotonidae), 达乌尔鼠兔。
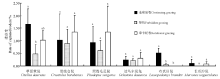
2.1.1 不同放牧方式下啮齿动物捕获率 3个年度不同放牧方式下, 啮齿动物捕获率最高为连续放牧区(4.45%), 其次为季节轮牧区(4.09%), 禁牧区捕获率最低(2.22%)(图2)。连续放牧区捕获率显著高于禁牧区(P< 0.05), 季节轮牧区捕获率与其它两个放牧区差异不显著(P> 0.05)。草原黄鼠在连续放牧区的捕获率显著高于禁牧区(P< 0.05), 与季节轮牧区的差异不显著(P> 0.05); 黑线毛足鼠、黑线仓鼠、达乌尔鼠兔在季节轮牧区中的捕获率为最高, 但3种不同放牧方式下均无显著差异(P> 0.05); 布氏田鼠在季节轮牧区中无捕获, 其捕获率在连续放牧区显著高于禁牧区(P< 0.05); 长爪沙鼠在禁牧区中无捕获, 连续放牧区和季节轮牧区中的捕获率也较少(图3)。
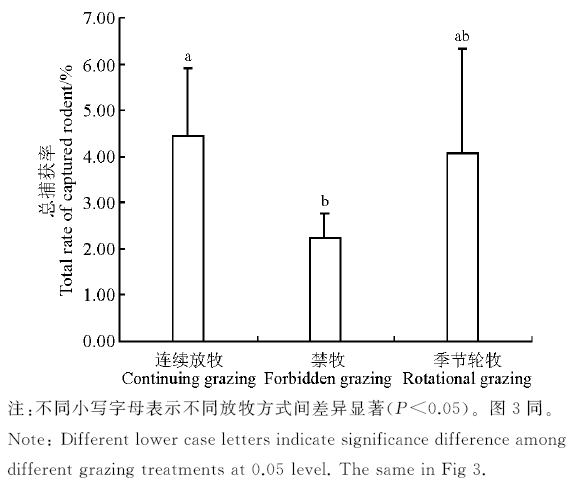 | 图2 不同放牧方式下的啮齿动物总捕获率Fig.2 Total rate of captured rodent in different grazing treatments |
 | 图3 不同放牧方式下个各鼠种的捕获率Fig.3 Rate of captured rodent in different grazing treatments |
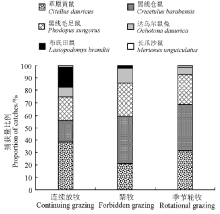
2.1.2 不同放牧方式下啮齿动物群落结构 本研究中不同放牧方式下的啮齿动物群落组成发生了一定变化(图4)。连续放牧区主要以草原黄鼠为主, 捕获量比例为38.60%; 季节轮牧区黑线仓鼠和草原黄鼠所占比例均较大, 捕获量比例分别为37.30%和31.50%; 禁牧区以黑线仓鼠捕获量比例最大, 为38.89%。
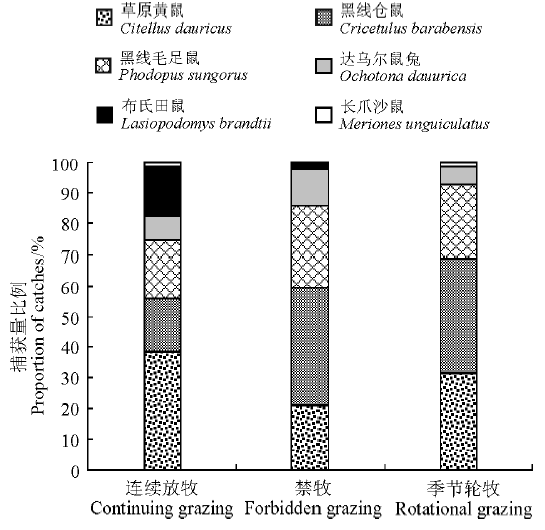 | 图4 不同放牧方式下啮齿动物群落结构Fig.4 Community structure in different treatments |
根据不同放牧方式下啮齿动物捕获情况, 分别计算了各放牧区啮齿动物群落丰富度指数、Shannon-Wiener指数、均匀性指数。结果表明, 连续放牧区的丰富度指数与Shannon-Wiener指数均最高, 且显著高于禁牧区(P< 0.05); 虽然季节轮牧区的啮齿动物群落均匀性指数较高, 但在3个放牧方式间无显著差异(P> 0.05); 禁牧区的丰富度指数、Shannon-Wiener指数及均匀性指数均为最低(表3)。
| 表3 不同放牧方式下啮齿动物群落多样性特征 Table 3 Diversity characteristics of rodent communities in different grazing treatments |
放牧通过改变植被高度、盖度、生物量和群落结构等影响啮齿动物食物资源[14]及隐蔽场所。在北美放牧对啮齿动物群落影响的研究中发现, 放牧增加了更格卢鼠(Dipodomys spectabilis)的数量[15]。付和平等[16]在阿拉善荒漠啮齿动物群落格局调查中发现, 三趾跳鼠(Dipus sagitta)对过度放牧生境表现出较强的适应性, 黑线仓鼠对郁闭度较高的禁牧生境表现出较强的适应性。刘书润[17]、董维惠等[18]在锡林郭勒典型草原布氏田鼠的研究中发现, 布氏田鼠的数量会在放牧导致的退化草地明显增加。武晓东和付和平[19]在不同放牧下啮齿动物敏感性研究中发现, 啮齿动物对不同放牧方式下的敏感性各不相同。草原黄鼠、三趾跳鼠更适应过度放牧区, 而黑线仓鼠则更适应禁牧区。刘伟等[20]研究表明, 放牧强度的增大引起适应隐蔽生境的根田鼠(Microtus oeconomus)数量减少。以上研究均表明, 不同啮齿动物对于放牧有着不同的响应。更格卢鼠、三趾跳鼠和布氏田鼠均属于体型中等以上物种, 它们适于在视野开阔的环境生存, 便于观察天敌从而降低捕食风险。而根田鼠和黑线仓鼠均为体型较小, 主要以种子为食。在郁闭度较高的环境中容易找到食物资源, 并且不容易暴露在天敌的视线当中, 因此其数量在放牧环境中有所减少。本研究表明, 连续放牧区植被高度明显低于禁牧样地, 因此, 适于开阔环境生存的鼠类, 其捕获率有所增加, 支持了前人研究结果, 即放牧增加了适于开阔环境生存的啮齿动物数量[15, 16, 17, 18, 19]。连续放牧不仅显著增加了草原黄鼠的数量而且还为布氏田鼠提供了适宜的生存条件, 从而导致局部啮齿动物种群数量的增加。禁牧中由于郁闭度高, 喜于开阔环境的啮齿动物不适于郁闭环境, 转移到其它适于生存的环境, 导致局部啮齿动物种群数量有所下降。
由于啮齿动物生物学及生态学特征的不同, 对于不同放牧方式下的响应也有所差异[21]。因此, 导致一些物种在放牧环境下数量增长, 而另一些物种在放牧环境下数量减少。因此, 这种啮齿动物群落数量、组成及结构的变化必然导致啮齿动物群落多样性的改变。国内外关于放牧与啮齿动物多样性的研究较多, 但其结果不尽相同。一些结论支持放牧降低啮齿动物群落多样性。如, 山地荒漠草原啮齿动物群落多样性及丰富度随着放牧强度的增加而减少[22]。半干旱荒漠草原未放牧区域的小型哺乳动物的物种丰富度和多样性明显大于放牧区物种丰富度和多样性[23]。而另一些研究结果支持放牧增加啮齿动物群落多样性。如, 放牧导致北美荒漠草原植被盖度低的地区啮齿动物群落多样性增高[24]。Rosi等[25]在阿根廷中部蒙特地区放牧研究中发现, 放牧导致了环境的异质性, 增加了对放牧环境有较高适应性的物种及其数量, 从而增加了啮齿动物多样性。本研究支持后一观点, 即放牧增加啮齿动物群落多样性。丰富度指数、Shannon-Wiener指数、均匀性指数最高值均出现在放牧区, 尤其是连续放牧区, 丰富度指数、Shannon-Wiener指数均显著高于禁牧区的(P< 0.05)。群落多样性指数的大小不仅与生境条件的差异有一定的关系[26], 而且与群落内各种群数量的均衡分配相关。不同物种对于放牧干扰的敏感性不同[19], 导致不同放牧方式下鼠类的分布发生了一定变化。连续放牧增加了适于开阔生境生存的物种草原黄鼠、布氏田鼠的数量从而增加了群落多样性。而禁牧区高的郁闭度导致适于开阔生境生存的物种慢慢迁出该地, 从而降低了物种丰富度。这与武晓东等[27]在阿拉善地区对不同放牧条件下, 禁牧啮齿动物物种数为最少的研究结果一致。
综上所述, 连续放牧会导致啮齿动物群落多样性增加, 但也因为草地的退化为一些鼠种提供有利的生存条件, 使得某些鼠种数量增长。禁牧区啮齿动物的数量虽然保持在最低水平, 但多样性也最低, 因此, 禁牧可能会减少种群间的竞争力从而促使某单一鼠种爆发提供有利条件, 从而增加鼠害发生的可能性。只有季节轮牧当中多样性指数及捕获率均保持在中等水平上, 因此, 本研究认为适当的放牧有利于维持啮齿动物多样性并且可以有效控制啮齿动物种群数量, 在等放牧面积中季节轮牧方式更利于合理利用草场资源并且能够抑制鼠害发生。
致谢:本研究工作得到了内蒙古呼伦贝尔盟草原工作站的大力支持; 内蒙古农业大学硕士研究生乌云嘎、卢志宏、岳闯、柴享贤, 包达尔罕等在野外数据搜集过程中给予了大力帮助, 在此一并表示感谢!
The authors have declared that no competing interests exist.
| [1] |
|
| [2] |
|
| [3] |
|
| [4] |
|
| [5] |
|
| [6] |
|
| [7] |
|
| [8] |
|
| [9] |
|
| [10] |
|
| [11] |
|
| [12] |
|
| [13] |
|
| [14] |
|
| [15] |
|
| [16] |
|
| [17] |
|
| [18] |
|
| [19] |
|
| [20] |
|
| [21] |
|
| [22] |
|
| [23] |
|
| [24] |
|
| [25] |
|
| [26] |
|
| [27] |
|