
{kind=link}
饲料添加剂植物多糖的生物学作用
[解玉怀1  , 尚庆辉
, 尚庆辉1 , 古丽美娜2 , 王英楠1 , 张崇玉1 , 杨维仁1 , 张桂国1 ]
, 尚庆辉, 张桂国]
|
|


第一作者:解玉怀(1991-),男,山东泰安人,在读硕士生,主要从事单胃动物营养的研究.E-mail:[email protected]
植物多糖是一类具有多种生物学功能的活性物质,作为功能性饲料添加剂被广泛应用于现代畜牧养殖业中,具有促生长,免疫增强,改善肠道健康,提高畜产品品质等多种生物学功能.本文综述了近年来在植物多糖生物学功能及作用机理相关领域的研究成果,并总结了其在畜禽养殖中的应用效果和存在的问题,为今后在此领域开展更广泛深入的研究提供参考.
, Shang Qing-hui, Zhang Gui-guo
Phytogenic polysaccharides are a class of active materials with various biological functions. They have been widely applied in livestock breeding as functional feed additives, which promote growth, strengthen immunity, improve bowel health and product quality. This paper summarized the research progress on the biological functions and working mechanisms of phytogenic polysaccharides, and explained the application effects as well as existing problems in livestock farming breeding, to provide a good reference for further exploration in this research area.
植物多糖是植物中存在的一类重要的生物活性物质, 是由10个以上单糖通过糖苷键聚合而成的一种天然生物大分子, 广泛存在于植物的根, 茎, 叶等组织器官中.枸杞(Lycium barbarum)多糖, 黄芪(Astragalus membranaceus)多糖, 牛膝(Achyranthes bidentata)多糖等近百种植物多糖已经被分离纯化, 并阐明了其部分生物学功能[1].研究发现, 水溶性植物源性多糖具有多种生物学功能, 具有在一定剂量范围内提高动物生长性能, 改善肠道环境, 促进消化道形态发育, 减少腹泻, 抗应激抗氧化, 增强免疫力, 防衰老等多种生物学功能[2, 3, 4].同时, 植物多糖作为一种多功能的天然植物成分, 具有来源广泛, 在畜禽体内及畜产品中无残留, 不产生抗药性等特点, 研究开发其用作新型饲料添加剂, 对于减少抗生素使用, 提高畜产品安全, 促进现代畜牧业的可持续发展具有重要的意义.
研究发现, 植物多糖的分子质量在几万至几百万道尔顿以上, 由一种或多种单糖残基按不同的比例以不同的糖苷键连接而成.组成多糖的单糖主要包括葡萄糖, 果糖, 阿拉伯糖, 木糖, 半乳糖, 鼠李糖, 岩藻糖, 糖醛酸, 甘露糖等.苜蓿草品种资源丰富[5], 其主要成分苜蓿(Medicago sativa)多糖是具有天然活性的植物多糖[6, 7], 由葡萄糖, 鼠李糖, 甘露糖, 半乳糖醛酸和另一种未知的单糖构成[8].由同一种单糖残基组成的多糖称为同质多糖, 如葡聚糖, 糖原, 直链淀粉, 纤维素等, 由不同种的单糖残基组成的多糖称为异质多糖, 如半纤维素, 果胶物质等.
与蛋白质相似, 多糖的结构也可分为一级, 二级, 三级和四级结构[9].多糖的一级结构是高级结构的基础, 包括所含单糖种类, 糖苷键类型, 有无分支, 分支的位置以及所含基团等.常见的糖苷键有α -1, 4-, α -1, 6-, β -1, 3-和β -1, 4-糖苷键, 单糖间连成直链, 也可以形成支链; 直链多以α -1, 4-和β -1, 4-糖苷键相连, 支链中链与链的连接点常为α -1, 6-糖苷键[10].研究表明, 多数具有突出活性的多糖都以-1, 3-糖苷键相连, 并以β -1, 3-D-葡聚糖的构成为主链, 同时具有一定的分支和特殊的基团[11].二, 三, 四级结构则分别涉及多糖分子主链的构象, 空间构象和非共价键形成的聚集体[12].
植物多糖生理作用广泛, 特别是在免疫调节方面, 可增强消化道黏膜免疫, 具有抗氧化, 抗病毒, 抗肿瘤等功能.在畜禽日粮中添加植物多糖, 可提高畜禽的生产性能, 并可提高营养物质的消化率[13, 14].人们对植物多糖及其复合物在畜禽上抗疾病, 促生长等方面的作用也有了越来越深入的认识.
在饲料中添加一定浓度的植物多糖能够改善动物的生产性能, 提高畜禽的平均日采食量和平均日增重, 降低料肉比.研究表明, 肉仔鸡日粮中添加1.0%和1.5%的水溶性苜蓿多糖(WSAP)能极显著(P< 0.01)提高42 d时的平均日增重和平均日采食量, 显著(P< 0.05)降低料肉比和腹脂率[15]; 刺五加(Acanthopanax senticosus)多糖在断奶仔猪上的应用也显示, 800 mg· kg-1的刺五加多糖可显著提高断奶仔猪的日增重, 降低料肉比和腹泻率[13].
研究表明, 植物多糖主要通过促进机体生长相关激素的合成与分泌来促进脂类与糖的代谢, 提高机体氨基酸与蛋白质合成, 以达到促生长的作用.机体的生长由生长激素轴来调控, 生长激素释放因子(GHRH), 生长激素(GH)和胰岛素样生长因子(IGFs)构成了调控轴.GH-IGF-I轴是生长激素轴的主要部分, 与机体蛋白, 脂类和糖的代谢密切相关, 其中GH是调控整个机体生长最主要的激素[16], 对各种细胞的代谢调节过程, 以及促进氨基酸的转运和刺激蛋白质的合成有重要的作用[17]; IGF-I能够对所有胰岛素靶器官起到经典的胰岛素效应, 促进脂肪和糖元合成.植物多糖能够调节GH-IGF-I轴上激素的分泌, 对提高动物生产性能, 改善胴体组成, 提高肉质和饲料转化率起到一定作用.WSAP能够显著或极显著提高GH和IGF-I基因在肾脏, 肝脏和脂肪组织中的表达, 且随着WSPA添加水平的增多, 肝脏和脂肪组织中两者基因表达量也随之增多[15, 18].香菇(Lentinus edodes)多糖在饲料中应用也能够显著提高自主ADFI, 降低腹泻率, 提高GH水平[19].
甲状腺激素(T3, T4)为机体重要的代谢调节激素, 通过调节戊糖磷酸循环, 调控蛋白质, 糖原和脂肪的合成与分解, 为机体供给能量, 促进机体生长[20].此外, T3, T4能够控制机体GH基因的表达与合成, 调控IGF-I的DNA的翻译和转录过程[21].香菇多糖可提高仔猪T3, T4水平, 同时能够降低仔猪血清Cor水平, 缓解应激[22](表1).
| 表1 植物多糖促生长作用及其机理 Table 1 The growth-promoting function of phytogenic polysaccharide and its mechanism |
机体在应激和病理状态时, 产生过多的活性氧(ROS), 羟基等强氧化自由基, 造成细胞膜, 蛋白质, DNA等的损伤, 甚至导致细胞死亡[23, 24]; 脂质过氧化反应过程会产生脂质过氧化物(LPO)和丙二醛(MDA)等毒性物质, 使机体遭受更大的损伤.植物多糖普遍具有抗氧化功能, 可提高超氧化物歧化酶(SOD)等氧化性自由基清除酶的活性, 加快ROS的清除[25, 26, 27], 减少对机体的损伤.
被乙醇损伤肝脏的小鼠饲喂枸杞多糖后, SOD, 还原型谷胱甘肽(GSH), 谷胱甘肽过氧化物酶(GSH-Px), 过氧化氢酶(CAT)含量上升, MDA水平下降, 肝脏抗氧化功能显著提高, 缓解了乙醇对机体的损伤[28].多糖对SOD的提高加快了H2O2的形成以消除ROS; GSH-Px可将H2O2等过氧化物分解成无毒的物质, 并能通过细胞膜LPO阻断脂质过氧化链式反应; CAT也可催化H2O2释放氧以氧化某些酚类和胺类物质, 避免H2O2在体内的蓄积; GSH是体内重要还原剂, 避免蛋白, 核酸等物质被氧化.
云芝(Coriolus versicolor)多糖具有提高机体抗氧化性能的作用.给小鼠灌注云芝多糖后, 肝内还原型与氧化型谷胱甘肽比值下降[29].对局部缺血的小鼠使用当归(Angelica sinensis)多糖处理, 发现心脏组织和血清中SOD, CAT, GSH和GSH-Px水平上升, 抗氧化活性增强[30].
植物多糖抗氧化作用的机理研究包括:1)多糖直接作用于ROS, 通过快速摄取
肠道是营养物质消化吸收的主要场所, 其形态发育的成熟程度是肠道发挥其功能的基础.植物多糖能够改善肠道形态, 提高肠绒毛与隐窝深度之比, 增强肠道消化吸收的能力.如, 黄芪多糖可改善肉仔鸡肠道形态, 增加绒毛高度和粘膜厚度[33]; 在日粮中添加5%菊苣果糖或其它非淀粉多糖可以显著增加大鼠回肠绒毛高度[34].
肠道绒毛是机体营养物质吸收的主要部位, 肠隐窝是绒毛根部下陷至固有层而形成的管状结构, 绒毛高度和隐窝深度是小肠功能状态的反映, 两者的比值越高说明肠道的吸收能力越强; 反之, 越低.幼龄动物在断奶时, 由于受到饲料形态, 营养成分等变化的应激, 机体肠道组织结构受到破坏, 使得机体肠绒毛高度与隐窝深度的比值显著下降[35], 腹泻率提高.有报道发现, 植物多糖能够显著提高小肠绒毛高度与隐窝深度的比值, 在断奶仔猪日粮中添加0.3%白术(Atractylodes macrocephala), 黄柏(Phellodendron amurense), 党参(Codonopsis pilosula)等7种植物多糖的混合物后, 该比值分别比空白对照组和抗生素组高出11.99%和11.41%[36].
研究表明, 植物多糖减轻应激对肠道损伤, 促进肠道绒毛发育的机理可能是植物多糖通过提高肠道粘膜免疫以阻止病原体对肠道粘膜的破坏, 间接促进了肠道形态的发育[37, 38].红色微藻细胞的硫酸化多糖, 果胶或纤维素能够显著增加大鼠小肠和结肠的长度, 同时多糖也增加了粘膜和空肠肌层截面面积[39].
与消化道内环境相关的指标包括肠道pH, 消化酶种类, 活力, 相关的微生物菌群种类, 数量等, 这是维持肠道正常发育和良好消化吸收功能的重要条件.肠道pH是影响消化酶活性, 微生物生存和繁殖的基础, pH值的升高易造成消化酶活性降低, 病原微生物的增殖, 从而使得饲料消化率降低, 甚至导致腹泻等疾病[40].低聚果糖能够降低肠道pH, 抑制大多数革兰氏阴性菌在内的致病菌的生长[41].
植物多糖作用肠道环境的机理源于植物多糖一级构型中存在的独特的糖苷键结构.活性多糖多以-1, 2-或-1, 3-糖苷键相连, 以β -1, 2-或β -1, 3-糖苷键构成主链.但动物的α -淀粉酶无法分解-1-3-等β 型糖苷键, 只能水解α -1, 4-和α -1, 6-糖苷键.目前有报道表明, 口服的活性多糖能以大分子的形式吸收, 吸收的主要部位在小肠前段, 如六味地黄多糖[42], 右旋糖酐[43]等, 其吸收的机制目前认为可能与大分子物质蛋白酶, 胰岛素等相同, 即通过胞饮的方式进行吸收[44].同时, 未被吸收的多糖进入后段肠道后被微生物利用, 发酵降解为CO2和挥发性脂肪酸, 从而降低肠道pH.在影响消化道内环境的基础上, 又进一步影响到消化道内多种消化酶活性.消化酶是机体内催化各种生化反应的特殊蛋白质, 幼龄动物消化系统发育不成熟, 脂肪酶, 蛋白酶, 淀粉酶等内源酶分泌不足, 相关研究认为, 在肉仔鸡日粮中添加0.6%的黄芪多糖后, 胰腺蛋白酶和胰腺脂肪酶活性显著高于对照组, 各试验组的脂肪酶活性均极显著高于对照组[33].
肠道菌群的平衡是衡量动物机体肠道健康的另一个重要方面, 正常情况下, 肠道菌群与宿主的生长, 健康有着密切关系, 对生命活动的多种生理功能尤为重要[45].研究表明, 植物多糖能够有效降低断奶仔猪盲肠和结肠中大肠杆菌的数量, 减少腹泻的发生[46], 饲料中添加黄芪多糖也能够增加雏鸡肠道中乳酸菌和双歧杆菌数量, 减少大肠杆菌数量[47].
多糖调节肠道微生物生长的机理, 一方面源于胃肠道中不同微生物对植物多糖的不同利用情况.如低聚果糖为β -1, 2-糖苷键型, 肠道中乳酸杆菌和双歧杆菌能够利用低聚果糖合成K族, B族维生素和蛋白质供机体使用, 还可分解成有机酸, 使肠道pH降低, 抑制革兰氏阴性菌等大多数致病菌的生长[41], 由此提高肠道有益菌增殖来抑制致病菌生长, 改善肠道菌群状况, 促使动物健康生长.另一方面被证实是植物多糖改变了肠道粘膜免疫的结果[48].肠黏膜固有层中的浆细胞可分泌一种蛋白为IgA, 为机体分泌型免疫球蛋白(SIgA).SIgA是消化道黏膜免疫的重要环节之一, 是主要效应因子, 对肠道细菌的定殖程度和微生物在体内的平衡起到调控作用.SIgA能够干扰病原微生物与黏膜上皮细胞受体结合, 减少粘膜细胞表面病原体附着, 降低病原菌毒力, 影响免疫排异, 阻止病原体传播和进一步的感染, 因而在免疫防御中起着非常重要的作用[37, 49].研究表明, 黄芪多糖可以显著提高雏鸡肠道SIgA的分泌水平[50](表2).
| 表2 植物多糖改善肠道环境机理 Table 2 The mechanism of improving intestinal environment of phytogenic polysaccharide |
大量的研究发现, 植物多糖是重要的免疫调节剂, 能够激活免疫细胞, 提高机体免疫功能, 对机体特异性和非特异性免疫功能均具有增强作用.其作用机制是通过激活各免疫细胞(淋巴细胞, 巨噬细胞等)对细胞因子的释放, 调动补体系统, 促进抗体生成等功能, 从而实现多功能, 多层次的免疫调节作用[51].
巨噬细胞是机体免疫系统的重要成员, 由单核细胞衍化而成, 与中性粒细胞等协同抵抗病原体入侵, 清除衰老, 损伤, 死亡及突变的细胞及其代谢废物, 是机体免疫功能的基础.研究表明, 巨噬细胞表面有多种跨膜蛋白识别受体, 天然植物多糖主要通过结合细胞表面Toll样受体, CD14和CR3受体, MR或SR等受体进入细胞, 并主要通过活化丝裂原活化蛋白激酶(MAPKs)或核因子kB(NF-kB)而激活下游两条不同的信号转导途径, 从而刺激细胞因子IL-1β , IL-6以及TNF-α 等细胞因子的释放, 以达到增强巨噬细胞的免疫活性[52].
一氧化氮(Nitric Oxide, NO)是体内重要的细胞信使因子, 广泛参与机体免疫应答等多种生理和病理过程.当巨噬细胞被活化时, 释放的大量NO具有细胞毒性, 可杀伤病原微生物, 也可激发炎症反应保护机体.盐角草(Salicornia herbacea)多糖能够提高巨噬细胞RAW264.7和小鼠腹腔巨噬细胞释放NO的活性, 亦可促进巨噬细胞释放细胞因子[53].Schepetkin等[51]报道, 从落基山桧(Juniperus scopolorum)中分离的水溶性多糖可显著提高巨噬细胞J774.A1中TNF-α , IL-6, IL-12, MCP等促炎症因子和IL-10抗炎症因子的产生.
淋巴细胞是机体重要免疫防御细胞, 天然活性多糖可以通过结合T细胞表面TCR/CD3受体, 并通过PTK介导下游的MAPKs和PKC/PLCγ 两条信号通路进行免疫应答的传导; 对于B淋巴细胞, 多糖主要通过结合细胞表面TLR2/4或IgM/CD79受体, 并介导下游MAPKs或NF-kB两条通路进行细胞信号的传导[54](图1).
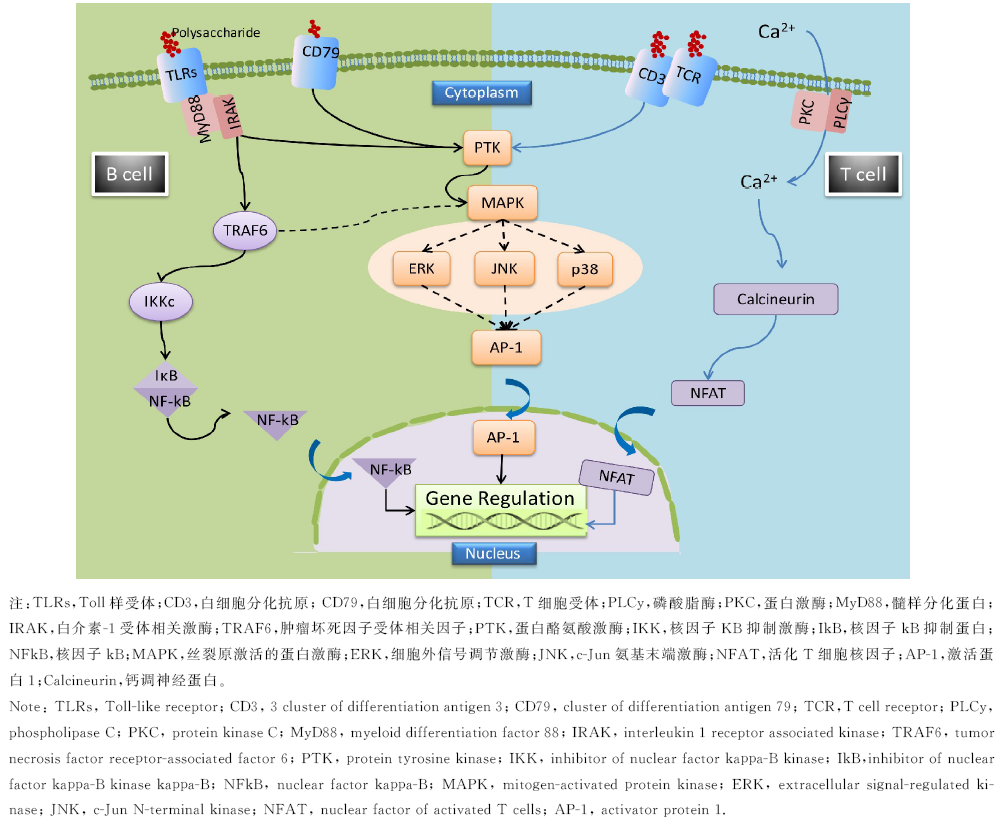 | 图1 植物多糖激活T, B淋巴细胞主要信号通路[54]Fig.1 The key signaling pathways of T, B lymphocyte activated by phytogenic polysaccharide[54] |
Chen等[55]研究发现, 枸杞多糖能够激活转录因子NFAT(Nuclearfactor of Activated T Cells)和AP-1, 促进T细胞跨膜蛋白CD25的表达, 并诱导IL-2和IFN-γ 基因的转录和蛋白质合成, 显著提高T淋巴细胞的增值活性.灵芝多糖可以促进小鼠脾细胞TNF-α , IFN-γ , IL-1, IL-6等细胞因子的分泌, 同时研究证实, 灵芝(Ganoderma Lucidum)多糖是B淋巴细胞的选择性刺激性多糖, 能够增加B细胞表面CD71+/CD25+和IgM的表达, 诱导小鼠脾淋巴细胞增殖与向IgM分泌型浆细胞的分化[56, 57].牛膝多糖, 白术多糖和黄芪多糖可以显著提高断奶仔猪外周血淋巴细胞转化率, 显著提高血清细胞因子IL-2, IL-1β , TNF, IFN-γ 的分泌量[58].
补体系统是存在于机体血清与组织液中经活化后具有酶活性的蛋白质, 可补充和辅助特异性抗体, 介导免疫溶血和溶菌作用.Yamada等[59]发现魁蒿(Artemisia princeps)多糖去除乙酰基后, 激活补体能力大大提高.
前人研究表明, 各种天然植物活性多糖均可发挥促生长, 抗氧化, 免疫调节和促肠道发育等生物学功能外, 某些多糖在动物上还具有抗肿瘤, 抗辐射, 降血糖, 降血脂, 降胆固醇[60, 61]等作用, 不同植物来源的多糖, 其包含的单糖苷种类和比例及后期的高级结构形态都有很大差别, 此外, 一些多糖分子中还具有特殊的糖配基, 这种组成和机构的差异可能是造成不同来源多糖主要生物学功能差异的主要原因.Chien等[62]发现, 灵芝多糖的抗肿瘤作用与NK细胞的功能有着密切关系, 灵芝多糖能够通过提高NK细胞的活性来刺激IL-2, TNF-α 和IFN-γ 等细胞因子的释放, 从而起到抗肿瘤的作用.研究表明, 黄芪多糖可对糖尿病大鼠起到降血糖作用, 其主要作用机制是降低糖尿病大鼠血清中GH浓度, 升高IGF-I, 从而发挥降血糖的作用[63].
植物多糖作为一类具有重要生物活性的新型植物源性饲料添加剂, 其优点是来源广泛, 在动物体内及畜产品中无残留, 无毒副作用, 不易产生抗药性.随着研究的不断深入, 植物多糖所具有的抗氧化, 免疫增强, 促生长等生物学功能逐渐被认识并在生产中得到广泛的应用.但目前为止, 对于植物多糖是通过怎样的途径作用于靶器官而完成体内免疫调控等各生物学功能的作用机制还不清楚, 有待进一步研究阐明.同时多糖在激素调节, 促进肠道发育及微生物增殖等方面的机理研究尚停留在推测和分析阶段, 缺乏系统的科学依据与结论.未来的研究将围绕探索多糖的各种生物学功能机制而展开, 从细胞和分子水平, 阐明其体内作用的分子通道.对于多糖能否影响基因表达还需做进一步的深入探索, 这些方面的研究将为阐明植物多糖分子作用机制提供理论支持, 同时也会推动植物多糖在生产中更广泛应用.此外, 植物多糖的分离纯化和定性定量的检测, 也是需要深入探讨的课题.目前提取多糖常用的水溶-醇析法, 工艺较为复杂, 所用的实际价格偏高, 不利于植物多糖在生产中的推广应用.常用的Sevag法纯化中提高多糖得率的方法也需要进一步改善.
我国植物资源丰富, 不同来源的植物多糖开发利用的空间较大.随着对多糖分离纯化, 结构特性, 效应机理研究的不断深入, 植物多糖作为一类新型饲料添加剂将具有更加广阔的应用前景.
The authors have declared that no competing interests exist.
| [1] |
|
| [2] |
|
| [3] |
|
| [4] |
|
| [5] |
|
| [6] |
|
| [7] |
|
| [8] |
|
| [9] |
|
| [10] |
|
| [11] |
|
| [12] |
|
| [13] |
|
| [14] |
|
| [15] |
|
| [16] |
|
| [17] |
|
| [18] |
|
| [19] |
|
| [20] |
|
| [21] |
|
| [22] |
|
| [23] |
|
| [24] |
|
| [25] |
|
| [26] |
|
| [27] |
|
| [28] |
|
| [29] |
|
| [30] |
|
| [31] |
|
| [32] |
|
| [33] |
|
| [34] |
|
| [35] |
|
| [36] |
|
| [37] |
|
| [38] |
|
| [39] |
|
| [40] |
|
| [41] |
|
| [42] |
|
| [43] |
|
| [44] |
|
| [45] |
|
| [46] |
|
| [47] |
|
| [48] |
|
| [49] |
|
| [50] |
|
| [51] |
|
| [52] |
|
| [53] |
|
| [54] |
|
| [55] |
|
| [56] |
|
| [57] |
|
| [58] |
|
| [59] |
|
| [60] |
|
| [61] |
|
| [62] |
|
| [63] |
|