
{kind=link}
{kind=link}
模拟氮沉降对三江平原小叶章湿地土壤细菌多样性的影响
[隋心1, 2  , 张荣涛
, 张荣涛2 , 杨立宾2 , 许楠2 , 柴春荣2 , 王继丰2 , 付晓玲2 , 钟海秀2 , 邢军会2 , 张玉2 , 倪红伟2 ]
, 张荣涛]
|
|


第一作者:隋心(1982-),男,黑龙江拜泉人,研究实习员,博士,研究方向为生态学。E-mail:[email protected]
利用黑龙江省科学院自然与生态研究所三江平原湿地生态定位研究站内的模拟氮沉降试验平台,应用Miseq测序技术对细菌16S rDNA进行序列测定和分析,探讨添加氮对三江平原小叶章湿地土壤细菌群落结构的影响。结果表明,与对照相比,添加氮增加土壤细菌群落多样性,低氮(4 g·hm-2·a-1)时土壤细菌多样性最高,高氮(8 g·hm-2·a-1)次之。模拟氮沉降导致湿地土壤细菌群落的结构以及丰度发生变化,低氮、高氮和对照地的优势种群均为酸杆菌门(Acidobacteria),变形菌门(Proteobacteria)次之,随着氮沉降的增加,酸杆菌门的丰度逐渐下降,变形菌门的丰度则先升高后降低。外源氮的输入使得土壤的理化性质发生了改变,导致细菌群落的组成以及细菌的相对丰度发生改变。16S rDNA heatmap分析则表明,不同的细菌群落对氮沉降的响应不同。本研究为预测未来大气氮沉降对三江平原湿地土壤微生物的影响以及湿地生态系统的变化提供基础数据和理论参考。
, Zhang Rong-tao
Wetland soil bacterial community structure under the simulation nitrogen depositions was studied using 16S rDNA gene sequencing. The soil samples were collected from three different nitrogen depositions in the Sanjiang Plain Field Experiment Station of the Institute of Nature & Ecology, Heilongjiang Academy Sciences. Total microbial DNA was directly extracted from soil samples and 16S rDNA genes were sequenced and analyzed by Miseq molecular technology. The results showed that both low N and high N treatment increased the bacterial diversity compared with control treatment and low N treatment had the highest diversity. The wetland bacterial community structures significantly changed under simulation N depositions. In low N treatment, high N treatment and control treatment, Acidobacteria were predominated followed by Proteobacteria. The richness of Acidobacteria decreased and the richness of Proteobacteria increased first then decreased along the deposition. Soil pH decreased with deposition, which showing acidic property and increasing the Acidobacteria distribution. Soil physicochemical properties changed by input N which resulted in the soil bacterial community composition and abundant changed. The heatmap tree based on the 16S rDNA sequences showed that different bacterial community structure differently responded on Nitrogen deposition. This research provided basic data and theory consultation for predicting the influence of future atmospheric nitrogen deposition on soil microorganism in Sanjiang Plain wetlands.
近几十年来, 由于科技进步以及工业的高速发展, 燃烧大量的化石燃料导致了大气氮沉降量迅速增加。根据欧洲NITREX(Nitrogen Saturation Experiments)项目计算, 森林每年氮饱和的临界负荷的最小值为10 kg· hm-2[1], 但是到目前为止, 中欧森林每年的大气氮沉降量达到了25~60 kg· hm-2, 已经严重超出了森林的氮饱和的最小值[2]。从全球来看, 在北美洲有些森林每年的大气氮沉降量也超出了森林的氮饱和的最小值, 达到40 kg· hm-2[3, 4]。已有研究表明, 我国降水中NO3-离子的浓度(3.2× 10-7 mol· L-1)和沉降量(2.0× 10-7 mol· L-1)与美国和日本接近, 但是我国降水中的NH4+离子的浓度则更高, 其沉降量也更大, 离子浓度高达3.77× 10-6 mol· L-1, 分别是美国和日本的4倍和3.7倍, 导致我国氮沉降量十分巨大, 分别是美国和日本的3倍和1.8倍[5]。目前, 大气氮沉降已经成为影响全球变化的重要现象之一, 受到了全世界的关注。近几十年来, 由于大气氮沉降的沉降量不断增加, 给全球生态系统造成了严重的影响, 例如土壤酸化的加重, 生物功能和多样性的降低等[6, 7, 8], 引起了全世界的广泛关注[9, 10]。
近年来, 随着分子生物学技术的发展, 环境微生物的研究方法也从以前的实验室纯培养进入到了分子生物学方法阶段。Handelsman等[11]在1998提出宏基因组(Metagenome)的概念, 它包括环境中所有微生物的基因, 目前主要指细菌和真菌的基因组总和。宏基因组学(Metagenomics)作为分子生态学的一种典型技术策略, 能够从环境样品中直接获得的基因组中进行文库构建、克隆、测序[12], 可以从群落水平上了解微生物完整的群落结构, 并且真实地揭示环境微生物群落的多样性和复杂性[13], 在环境微生物群落结构的研究中显示出无比的优越性[14], 在土壤、水体、湿地以及极端环境微生物多样性等研究方面被广泛应用[15, 16, 17, 18]。
三江平原湿地是我国面积最大、分布最为集中的湿地[19], 由于人类活动的影响, 湿地遭到破坏, 面积大幅减少, 已经从1949年的534.5万 hm2减少到148.16万hm-2[9], 湿地生态系统结构与功能发生了退化。目前研究主要集中在植物群落变化[10]、温室气体排放[19]和营养物质循环[20]。国外大量的研究已经表明, 氮沉降长期增加会对土壤微生物产生不利影响[21, 22, 23, 24, 25], 但是有关氮沉降对三江平原小叶章(Deyeuxia angustifolia)湿地的土壤微生物的结构组成和变化的影响, 到目前为止还没有相关的研究。因此, 本研究通过野外原位试验, 探讨模拟氮沉降对三江平原小叶章湿地土壤细菌多样性的影响, 以期为研究三江平原湿地生态系统在氮沉降背景下的变化提供科学依据, 并为三江平原湿地的保护与管理提供理论参考。
研究样点设置在黑龙江省科学院自然与生态研究所三江平原湿地生态定位研究站— — 洪河国家级自然保护区内(47° 45'39″ N, 133° 37'04″ E)。研究站海拔55~65 m, 多年平均气温1.9 ℃, ≥ 10 ℃有效积温2 165~2 624 ℃· d。多年平均降水量为585 mm, 50%~70%集中在7-9月, 多年平均蒸发量为1 166 mm。主要植被类型为草甸和沼泽, 优势植物有小叶章、狭叶甜茅(Glyceriaspiculosa)、毛果苔草(Carex lasiocarpa)、漂筏苔草(Carex pseudocuraica)等[26]。
2010 年5 月在研究区设置9块20 m× 20 m的小叶章沼泽化草甸湿地, 设3个氮沉降梯度, 分别为对照N1 (0 g· hm-2· a-1)、低氮N2(4 g· hm-2· a-1) 和高氮N3(8 g· hm-2· a-1), 每种处理重复3次。每年5月份进行施氮处理, 将NH4NO3溶于水, 用花洒均匀喷施。于2014年6月采集土壤样品, 采取随机取样的方式, 在每个试验区分别用直径为4 cm的土钻按混合采样法采集0-10 cm 表层土, 每个土样有5~8 个采集点的土壤混合, 每个处理分别混合成一袋土, 共计3袋, 去除土样中的植物和动物残体等杂质, 混合均匀后用四分法取适量于自封袋中, 迅速带回实验室。取一部分保存在-80 ℃冰箱, 用于分析土壤细菌群落结构。另一部分风干、磨细过0.15 mm筛、1.00 mm筛, 用于测定土壤理化性质。
土壤有机碳利用碳氮分析仪测定(耶拿-2100S, 德国); 土壤中全氮采用半微量凯氏定氮法测定; N
土壤微生物总DNA提取采用美国MO BIO的强力土壤DNA提取试剂盒(PowerSoil® DNA Isolation Kit), 按照说明书的操作步骤进行提取。将提取出的土壤微生物总DNA溶解在100 μ L去离子水中, 取5 μ L的DNA用1.0%的琼脂糖凝胶电泳检测(0.5× TAE缓冲液), 分析DNA的完整性和相对浓度。
由上海美吉生物医药科技有限公司进行Miseq高通量测序。细菌16S rDNA扩增引物采用通用引物(8F/533R)。
1.6.1 有效序列和优化序列数据统计 由于采用多个样品平行测序的方法, 所以各样品中的序列均引入了一段标示其样本来源信息的barcode标签序列及前引物(Forward Primer)序列。本次分析根据barcode标签序列和前引物序列筛选出有效序列后, 将测序接头与barcode序列去除, 并对处理后的有效序列进行数据统计。
为了分析结果更精确, 需要对有效序列进行去杂。去掉小于150 bp和含有模糊碱基以及引物碱基对错配达到两个以上的序列, 从而得到优化后的序列, 用优化后的序列来进行后续生物信息分析。
1.6.2 OTU-based 分析 对所有样品进行划分可操作分类单元(Operationaltaxonomic unit, OTU ), 并对OTU进行生物信息统计分析。将优化序列选取长度大于350 bp的序列并截齐后与silva数据库中的微生物基因序列进行比对后对序列进行聚类。聚类分析使用的软件为Mothur和Chopseq(http://www.mothur.org/wiki/Main-Page)。
1.6.3 细菌群落多样性分析 细菌群落物种的丰富度(Richness)和多样性(Diversity)分别用Chao1和Shannon指数表示, 测序深度指数用覆盖度(Coverage)表示。计算公式如下:
Chao1=OTUs+n1(n1-1)/2(n2+1).
式中, OTUs=实际观测到的OTU数目; n1=只含有一条序列OUT数目; n2=只含有两条序列的OUT数目。
Shannon index(H)=-∑ Pi× log2Pi.
式中, Pi=ni/N, 表明第i个物种的相对多度; ni=含有i条序列的OTU数目; N=所有序列数。
Coverage=1-n1/N .
式中, n1=只含有一条序列的OTU数目, N=总序列数。
群落多样性分析使用的软件为Mothur (version v.1.30.1) 。
1.6.4 群落结构分析
将优化序列与silva库中的已知序列进行比对, 对OTU序列进行种属鉴定。在门(Phylum)的水平上做样品群落分布柱状图, 比较3个土样中的细菌群落分布情况。
模拟氮沉降样地的土壤理化性质分析结果显示(表1), 样地的土壤类型主要为草甸土, 且不同处理间的pH值存在差异, 其中高氮样地的pH均值最低(P< 0.05)。不同处理间土壤的全氮、硝态氮、铵态氮和速效磷存在显著差异(P< 0.05)。有机碳、全氮、硝态氮、全磷和铵态氮含量呈现随着氮沉降的增加而升高的趋势, 而速效磷含量则呈现随着氮沉降的增加而下降的趋势。
| 表1 模拟氮沉降条件下的土壤理化性质的比较 Table 1 The physical and chemical properties under different nitrogen concentration conditions |
通过Miseq高通量测序并优化后, 3个处理共获得39 979条序列, 总碱基数为15 845 087 bp, 平均碱基长度为396.32 bp, 其中301~400 bp长度的序列占总序列数的99.74%(表2)。
| 表2 16s rDNA有效序列的数量及分布 Table 2 The number and distribution of valid sequences of 16s rDNA |
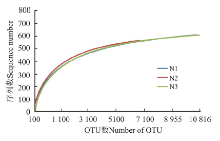
利用测序获得的序列进行随机抽样, 将抽到的序列数所代表的OTU数目构建稀释性曲线。3个处理的稀释性曲线均趋于平坦, 表明测序数据接近饱和, 测序的深度比较合理, 增加测序数量对发现新的OTU贡献率较小(图1)。
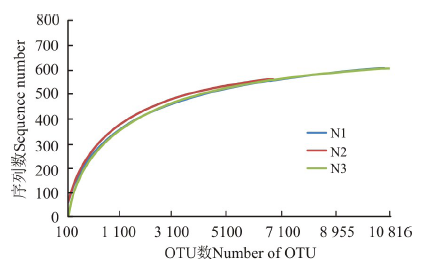 | 图1 不同处理土壤细菌OTU稀释曲线Fig.1 OTU rarefaction curves of soil bacteria in different treatments |
通过比对细菌16S rDNA的相似性, 将相似性高于98%的序列归为同一个OTU[27]。对不同氮沉降梯度下小叶章湿地土壤细菌16S rDNA多样性指数进行分析(表3), N1、N2和N3的Shannon指数分别为5.26、5.38和5.32, 丰富度指数Chaol分别为745、698和728。与N1相比, 随着氮浓度的逐渐增加, 细菌的多样性有所增多, 在N2时细菌多样性达到最高; 而细菌的丰富度指数与多样性的变化不同, 在N2时细菌丰富度最低。这表明不同氮沉降梯度下小叶章湿地土壤细菌的结构和丰富度均存在着差异, 且不同氮沉降梯度下土壤细菌的群落结构变化规律不同。
| 表3 模拟氮沉降各梯度下小叶章湿地土壤样本中细菌丰富度和多样性指数 Table 3 Bacterial diversity indices of Deyeuxia angustifolia wetlands under simulation nitrogen deposition |
将测序所得16S rDNA序列在Genbank中进行比对, 得到不同氮沉降梯度下小叶章湿地细菌的分类(表4)。N1、N2和N3小叶章湿地优势种群均为酸杆菌门(Acidobacteria), 分别占总测序数的53.31%、45.64%和36.33%。变形菌门(Proteobacteria)分布仅次于酸杆菌门, 在N1、N2和N3中也有大量的分布, 分别占总序列数的21.60%、21.98%和18.20%。另外, 浮霉菌门(Planctomycetes)、绿弯菌门(Chloroflexi)和硝化螺旋菌门(Nitrospirae)在3个处理中的比例均超过3%, 也有大量的分布。
| 表4 模拟氮沉降条件下土壤细菌16S rDNA序列分析 Table 4 Sequence analysis of 16S rDNAunder different nitrogen concentration conditions |
不同氮沉降的湿地中土壤微生物还包括拟杆菌门(Bacteroidetes)、绿菌门(Chlorobi)、芽单胞菌门(Gemmatimonadetes)、迷踪菌门(Elusimicrobia)、螺旋体菌门(Spirochaetae)、疣微菌门(Verrucomicrobia)、厚壁菌门(Firmicutes)、蓝藻菌门(Cyanobacteria)、装甲菌门(Armatimonadetes)、热胞菌门(Thermotogae)和纤维杆菌门(Fibrobacteres), 它们在N1、N2和N3三个处理中分布均比较少, 各类细菌占总序列数的比例都小于3%。另外, N1、N2和N3三个处理中均有≥ 17%的细菌在Genbank中没有被明确的分类, 属于未分类的细菌。
与N1相比, 氮沉降对酸杆菌门数量分布的影响最大, 随着氮浓度的增加, 土壤中酸杆菌门呈现逐渐下降的趋势。氮浓度增加后, 与N1相比, N2和N3的酸杆菌门分别减少了7.67和16.98个百分点。变形菌门的变化规律稍有不同, 与N1相比, N2增加了0.38个百分点, N3降低了3.4个百分点。此外, 浮霉菌门、绿弯菌门和硝化螺旋菌门受氮沉降的影响也比较大。与N1相比, 土壤氮含量增加后, N2的浮霉菌门增加了0.72个百分点, 而N3下降了0.83个百分点; 而N2和N3的绿弯菌门分别增加了4.64和2.84个百分点; 硝化螺旋菌门则在N2中几乎没有变化, 在N3中减少了0.49个百分点。
在属的水平上对各处理所含菌属进行聚类, 根据聚类后各处理中的不同OTU所含序列的丰度制作Heatmap(图2), 该图能够反映在菌属水平上各样品细菌群落结构与丰度上的差异。不同氮沉降梯度下湿地土壤16S rDNA细菌可以划分为5个聚类(Cluster)。菌属的丰度受到不同氮浓度的影响, 在所测得的菌属中丰度受不同氮浓度影响比较大的是链霉菌(Streptomyces)、Oryzihumus和嗜酸链霉菌属(Streptacidiphilus)。
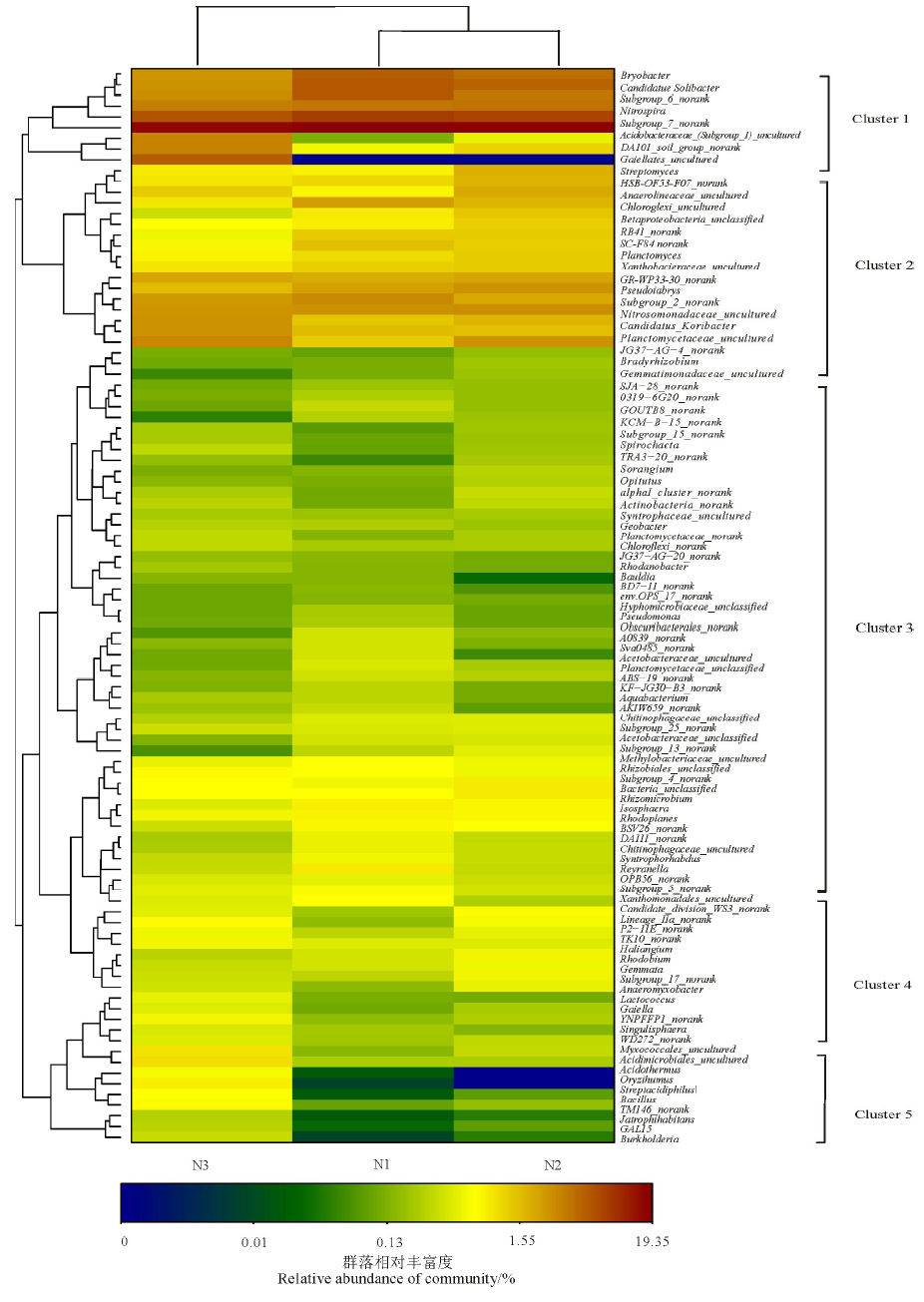 | 图2 基于细菌16S rDNA序列构建的热图Fig.2 Heatmap tree based on the bacterial 16S rDNA sequences |
Cluster 1主要包括Bryobacter、Candidatus、硝化螺旋菌、链霉菌等(图2)。氮沉降增加使得细菌结构的分布发生了明显的影响。例如, Bryobacter和Candidatus在N1中要高于N2和N3; 链霉菌的丰度在N3中要高于N1和N2。
Cluster 2主要包括绿弯菌门和硝化螺旋菌门两大类群。其中绿弯菌门的丰度在N3中比较低, 在N1和N2中较高; 而硝化螺旋菌门在N3中比较高, 在N1和N2中较低。
Cluster 3主要包括螺旋体菌、放线菌、地杆菌、光合细菌、醋杆菌、Aquabacterium、Chitinophagaceae等。螺旋体菌在N3中的丰度较低, 在N1和N2中丰度较高; 放线菌和地杆菌则在N1中丰度较低, N2和N3中丰度较高; 光合细菌和醋杆菌则在3个氮沉降样地中丰度接近; Aquabacterium和Chitinophagaceae则在N1中丰度较高, 在N2和N3中丰度较低。
Cluster 4主要包括Isosphaera、Chitinophagaceae、Syntrophorhabdaceae、黄单胞菌等。Isosphaera和Chitinophagaceae在N1中丰度较高, N2和N3中丰度较低; Syntrophorhabdaceae和黄单胞菌在N1中的丰度较低, N2和N3中的丰度较高。
Cluster 5主要包括Singulisphaera、黏球菌、酸微菌等。Singulisphaera、黏球菌、酸微菌在N3中的丰度要大于N1和N2中。
从总体来看, N1和N2的细菌结构和丰度都比较接近, N3的土壤细菌群落结构和丰度变化比较大, 这说明N3对土壤细菌结构的影响比较明显, 这从图2可以明显看出来。另外, 对于N1和N2两个小叶章湿地来看, 二者的细菌群落结构和丰度都比较接近, 这说明在N2的条件下, 土壤细菌的结构并没有发生显著的改变。
土壤pH值是影响土壤微生物多样性的重要因素之一, pH值的变化对土壤的有机质分解与合成以及N、P、K营养元素的合成与转化有着重要的作用[28]。在不同氮沉降条件下, N1的pH值最高, 其次是N2, 但二者无显著差异(P> 0.05)。氮沉降的增加会对土壤养分产生影响。将N2和N3两种处理与N1相比, 氮沉降使土壤有机碳、氮等的含量均显著增加(表1), 这可能是氮输入能促进碳固定, 从而增加土壤有机碳含量, 这与Conant等[29]的观点一致。说明外源氮素的输入增加了湿地生态系统可利用氮素状况, 从而对湿地土壤有机碳含量产生了重要影响。另外, 外源氮素的增加还促进了土壤硝态氮和铵态氮含量增加, 促进了土壤氮的矿化率, 使得土壤矿质氮的含量也相应的增加, 从而使土壤全氮的含量得到显著增加, 这和大多数学者的研究结果一致, 表明土壤硝态氮和铵态氮在土壤氮饱和之前会随着氮输入量的增加而增加[30]。在南亚热带森林土壤有效氮含量对于模拟氮沉降的初期响应的研究中, 发现大气的氮沉降会使土壤中的速效氮含量增加, 而且氮处理水平越高、土壤速效氮含量增加越多[31]。
多样性指数对于分析不同土壤的微生物群落结构是非常有效的方法, 多样性指数越高则微生物群落多样性越高, 它由种类的丰富度及种类的均匀度两部分组成[32, 33]。本研究表明, 不同氮沉降条件下土壤细菌的群落结构发生了显著变化, Shannon指数呈现出N2> N3> N1的趋势(表2), 可见, 氮沉降会增加速效氮含量增加, 而且氮处理水平越高、土壤速效氮含量增加越多[31]。
多样性指数对于分析不同土壤的微生物群落结构是非常有效的方法, 多样性指数越高则微生物群落多样性越高, 它由种类的丰富度及种类的均匀度两部分组成[32, 33]。本研究结果表明, 不同氮沉降条件下土壤细菌的群落结构发生了显著的变化, Shannon指数呈现出N2> N3> N1的趋势(表2), 可见, 氮沉降会增加细菌的多样性, 低氮较高, 高氮略有下降, 说明氮沉降增加细菌多样性具有阈值限制性, 即氮浓度增加到一定阈值时, 细菌多样性则由升高转变为下降。这可能是由于氮沉降增加了土壤中的营养物质, 为细菌的生长提供了丰富的营养来源, 因此, 一定浓度的氮素会促进细菌的生长, 这与对杉木林土壤微生物[34]的研究结果一致。但是在高氮水平下, 氮增加改变了土壤中营养物质的有效性, 这导致了某些微生物定阈值时, 细菌多样性则由升高转变为下降。这可能是由于氮沉降增加了土壤中的营养物质, 为细菌的生长提供了丰富的营养来源, 因此, 一定浓度的氮素会促进细菌的生长, 这和刘彩霞等[34]的研究结果一致。但是在高氮水平下, 氮增加改变了土壤中营养物质的有效性, 这导致了某些微生物种群(如偏于利用植物残体的种群) 的生长, 但是同时又抑制了其它种群, 致使群落多样性指数的下降。总的来说, 氮沉降会对土壤细菌的群落结构具有一定促进作用。由此可以推断, 目前我国一些地区的高氮沉降势必会对土壤微生物群落产生影响[35, 36, 37]。
通过16S rDNA序列分析表明, N3、N2和N1的优势种群都是酸杆菌门, 分别占53.31%、45.64%和36.33%。很多研究都表明, 湿地土壤中主要的细菌群落是酸杆菌门和变形菌门。例如, 三江平原湿地土壤细菌主要是酸杆菌[38], 还有研究表明, 湿地土壤中主要优势细菌群落为变形菌[39, 40], 本研究得到相似结果, 表明在湿地土壤中, 酸杆菌门和变形菌门是最主要的两大类群。
氮沉降改变了土壤细菌群落组成, 酸杆菌门的丰度随着氮沉降的增加而下降, 而变形菌门随着氮沉降的增加先升高后降低, 这和Zhou等[41]的研究结果一致, 表明酸杆菌门与土壤氮含量呈负相关关系。对于变形菌门的丰度来说, 低氮处理时, 变形菌门丰度最高, 高氮时出现下降。有一些研究结果表明, 变形菌门在长期施氮处理中的占主要优势[42, 43]。而本研究发现, 在高氮处理时, 土壤变形菌门丰度会下降, 这可能是由于在高氮浓度时, 土壤氮含量已经过饱和, 过饱和氮反而会抑制土壤养分的供给, 导致变形菌丰度下降。
本研究结果还显示, 氮沉降导致土壤矿质氮含量大幅上升, 土壤呈现弱酸性, 而3个处理中都含有较多的酸杆菌。这可能是与弱酸性的土壤环境有关, 土壤的弱酸性会有利于某些酸杆菌的生长。湿地中土壤微生物在湿地土壤的物质循环和能量流动中起非常重要的作用。但目前对湿地的研究还相对较少, 对氮沉降的研究更为薄弱。这是因为湿地与森林、草原等生态系统相比, 由于其环境特殊, 季节性或常年积水, 所以在不同地点、不同季节的研究结果差别会较大。因此, 研究湿地土壤微生物是一个十分复杂和庞大的系统工程, 对其研究需要不断地从各个方面积累, 并应有多个学科的渗透与加入。全球大气氮沉降增加背景下我国东北地区的氮沉降将会增加。在此背景下, 研究湿地土壤细菌群落组成和结构对氮沉降的响应, 对研究未来气候变化对湿地生态系统多样性格局和功能稳定性的影响具有重要参考意义。
The authors have declared that no competing interests exist.
| [1] |
|
| [2] |
|
| [3] |
|
| [4] |
|
| [5] |
|
| [6] |
|
| [7] |
|
| [8] |
|
| [9] |
|
| [10] |
|
| [11] |
|
| [12] |
|
| [13] |
|
| [14] |
|
| [15] |
|
| [16] |
|
| [17] |
|
| [18] |
|
| [19] |
|
| [20] |
|
| [21] |
|
| [22] |
|
| [23] |
|
| [24] |
|
| [25] |
|
| [26] |
|
| [27] |
|
| [28] |
|
| [29] |
|
| [30] |
|
| [31] |
|
| [32] |
|
| [33] |
|
| [34] |
|
| [35] |
|
| [36] |
|
| [37] |
|
| [38] |
|
| [39] |
|
| [40] |
|
| [41] |
|
| [42] |
|
| [43] |
|