






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
兰州银滩黄河湿地4种植物的光合特性
[张华1, 2  , 康雅茸
, 康雅茸1 , 徐春华1 ]
, 康雅茸|
|


本研究以兰州银滩黄河湿地自然生长的千屈菜( Lythrum salicaria)、芦苇( Phragnites awstralis)、藨草( Scirpus triqueter)、菖蒲( Acorus calamus)为研究对象,对4种典型植物生长季不同阶段的光合特性及其与环境因子间的关系进行比较分析,探讨了4种植物对湿地的适应特性和机制。结果表明,1)5月11日千屈菜、芦苇的净光合速率(Pn)呈不明显的双峰曲线,藨草和菖蒲的Pn变化较为平稳,日均Pn表现为芦苇>千屈菜>菖蒲>藨草;8月19日,千屈菜、芦苇、藨草的Pn呈双峰曲线,光合“午休”现象明显,菖蒲为单峰曲线,日均Pn表现为芦苇>菖蒲>千屈菜>藨草。2)5月11日,光合有效辐射(PAR)、大气温度(Ta)、大气CO2浓度(Ca)、相对湿度(RH)、蒸腾速率(Tr)、气孔导度(Gs)、胞间CO2浓度(Ci)、水分利用效率(WUE)与4种植物Pn的日变化存在不显著、显著( P<0 .05)或极显著( P<0 .01, P<0.001)的正相关关系;8月19日,PAR、Ca、Tr、Gs、WUE、土壤湿度(SM)与4种植物Pn的日变化存在不显著、显著或极显著的正相关关系;3)5月11日,PAR、Ca、Ta、RH对4种植物的Pn影响较大,是主要的环境生理影响因子;8月19日,PAR、Tr对千屈菜、藨草的Pn影响最大,PAR、WUE对芦苇、菖蒲的Pn影响较大,是主要的环境生理影响因子。尽管各因子对4种植物的光合速率均有不同影响,但PAR是对植物光合生理特性产生影响的根本原因。
The present study analyzed the relationship between photosynthetic characteristics in different growth stages and environmental factors and discussed plant adaption characteristics and mechanism in wetlands based on natural plants of Lythrum salicaria, Phragmites australis, Scirpus triqueter and Acorus calamus’s growing in Yintan wetland of Lanzhou section of Yellow River. In May 11, the two peaks curve of net photosynthetic rate (Pn) of L. salicaria and P. australis was not obvious and the Pn of S. triqueter and A. calamus were relatively stable, the average of Pn decreased in the following order: P. australis>L. salicaria>S. triqueter>A. calamus. In August 19, the curves of Pn of L. salicaria, P. australis and S. triqueter showed two peaks with significant photosynthetic “midday depression” and the curves of Pn of A. calamus was one peak, the average of Pn decreased in the following order: P. australis>A. calamus>L. salicaria>S. triqueter. In May 11, photosynthetically active radiation (PAR), atmospheric temperature (Ta), atmospheric CO2 concentration (Ca), relative humidity (RH), transpiration rate (Tr), stomatal conductance (Gs), intercellular CO2 concentration (Ci) and water use efficiency (WUE) had positive correlation or significant correlation with the diurnal variation of Pn. In August 19, only PAR, Ca, Tr, Gs, WUE and soil moisture had positive correlation or significant correlation with the diurnal variation of Pn. In May 11, PAR and WUE had the greatest effects on Pn of 4 studied plants, which were the major environmental and physiological factors. In August 19, PAR and Tr had the greatest effect on Pn of L. salicaria and S. triqueter’s and PAR and WUE had the greatest effect on Pn of P. australis and A. calamus which were the major environmental and physiological factors. Although each factors had different effects, PAR was the main cause of plant photosynthetic physiological characteristics.
植物的光合作用是构成植物生产力的最重要因素, 陆地生态系统吸收固定能量、分配转化物质和水碳循环的基本环节, 也是地球陆地系统表层物质循环和能量交换的重要生理生态过程, 以及物质复杂的生物化学进程[1, 2, 3]。对晴天和多云天气状况下的芦苇光合作用日变化进行研究, 发现晴天芦苇(Phragmites australis)净光合速率的日变化有明显的光合“ 午休” 现象, 多云天芦苇的净光合速率主要受光强控制, 随光合有效辐射的变化而变化[4]。对水葱(Scirpus validus)、菖蒲(Acorus calamus)、鸢尾(Iris tecorum)、千屈菜(Lythrum salicaria)这4种湿地植物的光合特性进行研究, 发现它们的净光合速率日均值种间差异不显著, 并且净光合速率日变化与气孔导度、光合有效辐射、胞间CO2浓度、蒸腾速率有极显著或显著的相关关系, 气孔导度是影响光合速率的主要生理生态因子[5]。对黄土高原甘肃红豆草(Onobrychis viceiaefolia)、沙打旺(Astragalus adsurgens)、东方山羊豆(Galega officinalis)、多年生香豌豆(G. orientalls)4种豆科植物的光合日变化进行比较, 结果表明红豆草和沙打旺的净光合速率及蒸腾速率显著高于山羊豆和多年生香豌豆, 属于高光合、高蒸腾牧草, 4种豆科植物均表现出较高的水分利用效率及较低的水分消耗, 因此具有大面积推广的价值[6]。对塔克拉玛干6、7、9月份胡杨(Populus diversifolia schrenk)、疏叶骆驼刺(Alhagi sparsifolia)、头状沙拐枣(Colligonum caput-medusae)、多枝柽柳(Tamarix ramosissima)4种荒漠植物的气体交换与外界环境因子的关系做了比较研究, 结果表明, 环境因子中光照强度是最重要的主导因子, 其次是温度, 大气湿度与植物的蒸腾速率呈负相关, 荒漠植物的温度生态位会随气温的升高而增高, 并且它们都能调整叶温来适应沙漠地区的环境[7]。研究C3植物对环境因子的响应得出, 大多的C3植物气孔导度较高, 意味着更多的蒸腾失水将引起脱水的威胁[8]。对中东的沙拐枣进行了研究发现, 较高的光强有助于这种C4植物的光合作用[9]。国内外对旱生植物、湿生植物的研究较多, 但对半干旱地区湿地植物的研究较少, 不同月份的光合特性的比较研究却很少见。春季和夏季是西北地区植被的生长季, 光合作用主要在该期间进行, 为探讨黄河湿地典型植被生长季光合作用日变化特征及其影响因子, 本研究对千屈菜、芦苇、藨草和菖蒲4种植被优势种在不同生长季光合作用日变化特性进行对比研究, 分析生理与环境因子对4种湿地植物的影响程度, 以期为湿地植物的保护与培育提供参考。
研究区位于兰州黄河安宁段银滩大桥两侧黄河湿地内, 地理坐标为103° 42' E、36° 05'N, 海拔1 480.71 m。属中温带大陆性气候, 年平均温度9.3 ℃, 年平均降水量325 mm, 主要集中在7-9月, 占全年降水量的60%[10], 土壤以灰钙土为主, 成土母质为黄土[11], 河谷阶地多为次生黄土或原生黄土与次生黄土交错叠置, 质地疏松, 低洼地和季节性河床部分为沼泽土[10]。
千屈菜、芦苇、藨草(Scirpus triqueter)、菖蒲为黄河湿地的植被优势种, 均是多年生草本植物[12], 生命力旺盛, 喜光、喜湿, 是湿生植物。其中千屈菜、芦苇、菖蒲是C3植物, 藨草是C4植物[13, 14]。
本研究采用美国CI-340便携式光合测定系统和WatchDog 2000自动气象站, 于2014年5月和8月中旬晴朗无云的天气进行试验。在开放气路下对千屈菜、芦苇、藨草、菖蒲的功能叶片进行测定。每天08:00-18:00进行试验, 观测4种植物叶片的净光合速率(Pn)、光合有效辐射(PAR)、蒸腾速率(Tr)、气孔导度(Gs)、胞间CO2浓度(Ci)、叶面饱和水汽压差(VPD)。每隔1 h测定1轮, 每个叶片重复测定3次, 取平均值。待一天测量结束, 剪下所测植物叶片, 在室内进行拍照。同时观测试验地点的瞬时大气CO2浓度(Ca)、大气温度(Ta)、空气相对湿度(RH)、土壤温度(Ts)、土壤湿度(SM)。
在自然条件下, 强烈的太阳辐射是引起一天中温度、空气相对湿度等一系列环境条件变化的根本原因[17]。5月11日和8月19日的PAR从08:00开始增加, 分别在11:00和12:00达到最高, 为504和1 700 μ mol· m-2· s-1, 8月19日的日均PAR要比5月11日的高出704.97 μ mol· m-2· s-1(图1a)。大气温度也随着PAR的增加而增加, 5月11日08:00-13:00温度增长幅度为11.12 ℃(图1b), 8月19日08:00-13:00温度增长幅度为17.86 ℃, 两者相差较大, 5月11日大气温度在13:00达到峰值29.99 ℃, 8月19日在14:00达到峰值40.35 ℃, 之后温度逐渐降低, Ta在一定程度上会影响叶片的温度, 决定了光合作用的生化反应速率和VPD[18]。8月19日的大气CO2浓度明显高于5月11日的, 且两次测定均是08:00的CO2浓度最高(图1c), 然后逐渐降低。5月11日和8月19日间RH相差较小, 5月11日的RH略高于8月19日的空气RH, 且均在08:00时最高值, 随后也是逐渐降低(图1d)。
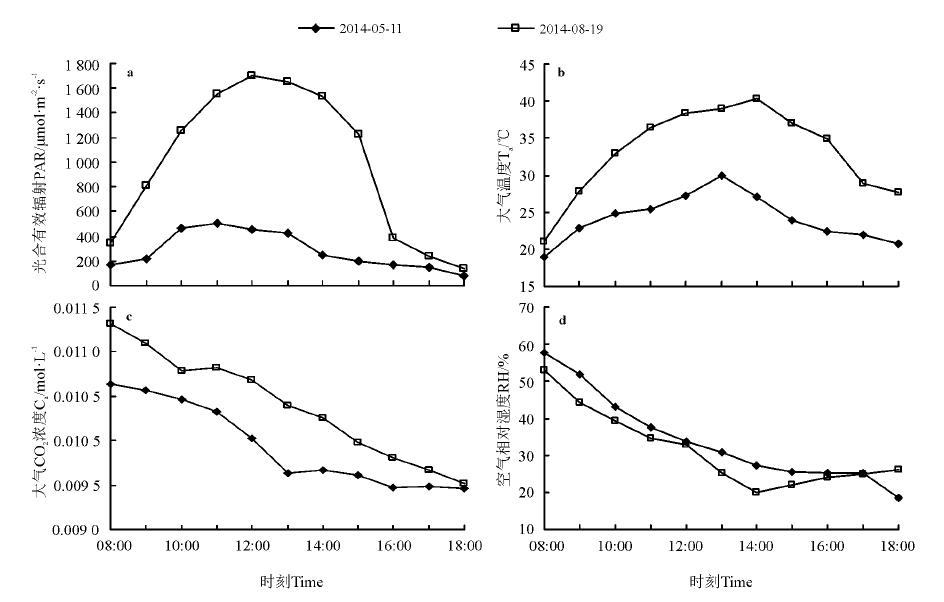 | 图1 银滩湿地环境因子的日变化Fig.1 Diurnal change of environmental factors in Yintan wetland |
5月11日千屈菜和芦苇的Pn具有明显的日变化, 均是轻微的双峰曲线, 在12:00达到第一峰值(0.039、0.099 μ mol· m-2· s-1), 第二峰值(0.022、0.043 μ mol· m-2· s-1)不明显, 藨草和菖蒲的Pn变化较为平稳。8月19日千屈菜、芦苇和藨草的Pn呈明显的双峰曲线, 千屈菜分别在11:00(0.042μ mol· m-2· s-1)、16:00(0.030 μ mol· m-2· s-1)点达到峰值, 芦苇分别在12:00(0.051 μ mol· m-2· s-1)、16:00(0.039 μ mol· m-2· s-1)达到峰值, 藨草的Pn变化较为平稳, 在11:00达到一个小峰值(0.013 μ mol· m-2· s-1), 菖蒲为单峰曲线在12:00达到峰值(0.047 μ mol· m-2· s-1)。5月11日日均Pn表现为芦苇> 千屈菜> 菖蒲> 藨草, 8月19日表现为芦苇> 菖蒲> 千屈菜> 藨草(图2)。
 | 图2 4种湿地植物种湿地植物净光合速率的日变化Fig.2 Diurnal net photosynthetic rate of 4 wetland plants |
5月11日千屈菜、芦苇、菖蒲的Tr均呈双峰曲线, 千屈菜在12:00以前呈上升趋势, 达到峰值(0.013 mmol· m-2· s-1)后然后波动变化; 芦苇在13:00以前呈上升趋势, 达到峰值(0.027 mmol· m-2· s-1)后迅速降低, 到15:00降到最低, 接着上升变化平稳; 藨草在13:00之前Tr的变化都较稳定, 14:00以后有所下降但变化较为稳定; 菖蒲在11:00以前呈上升趋势, 达到第一峰值(0.021 mmol· m-2· s-1)后迅速降低, 在15:00达到第二峰值(0.022 mmol· m-2· s-1), 接着波动下降。8月19日千屈菜、芦苇、藨草均呈现午间高峰的单峰曲线, 菖蒲为双峰曲线。5月11日和8月19日的日均Tr变化规律一致, 表现为芦苇> 菖蒲> 千屈菜> 藨草(图3)。
 | 图3 4种湿地植物蒸腾速率的日变化Fig.3 Diurnal of the transpiration rate of 4 wetland plants |
5月11日千屈菜的Gs在12:00以前呈现上升趋势, 12:00达到峰值后开始下降, 下降到一定程度则保持较为平稳的波动; 芦苇在13:00以前呈上升趋势, 13:00达到峰值后开始下降, 然后又缓慢上升; 藨草曲线变化较为平缓, 14:00降到最低值, 菖蒲呈“ V” 型曲线, 最低值出现在13:00。8月19日千屈菜、芦苇、菖蒲均表现为“ M” 型曲线, 藨草的变化较为平稳, 10:00以前呈上升趋势随后缓慢降低。 5月11日日均Gs表现为菖蒲> 芦苇> 千屈菜> 藨草, 8月19日表现为芦苇> 千屈菜> 菖蒲> 藨草(图4)。
 | 图4 4种湿地植物气孔导度的日变化Fig.4 Diurnal stomatal conductance of 4 wetland plants |
5月11日千屈菜Ci低谷与其Pn达到最大值的时刻一致(12:00), 芦苇、藨草、菖蒲Ci低谷出现时刻均滞后于最大Pn的出现时刻。8月19日4种湿地植物Ci低谷出现时刻也均滞后于最大Pn的出现时刻, 5月11日和8月19日4种植物的Ci总体变化趋势为早晚偏高, 中午低, 呈“ V” 型曲线(, 5月11日日均Ci表现为菖蒲> 千屈菜> 藨草> 芦苇, 8月19日表现为菖蒲> 藨草> 芦苇> 千屈菜(图5)。
 | 图5 4种植物胞间CO2浓度的日变化Fig.5 Diurnal intercellular carbon dioxide concentration of 4 wetland plants |
5月11日菖蒲的VPD最高, 变幅为2.065 kPa, 千屈菜、芦苇、藨草的变化曲线较一致, 8月19日4种植物较5月11日的变化幅度大, 芦苇的变幅最大, 为3.386 kPa。4种植物叶面VPD的日变化均为钟型曲线, 总体表现为先升后降, 菖蒲和芦苇的VPD均较高, 千屈菜、藨草的VPD均较低。5月11日日均VPD表现为菖蒲> 芦苇> 千屈菜> 藨草, 8月19日表现为芦苇> 菖蒲> 千屈菜> 藨草(图6)。
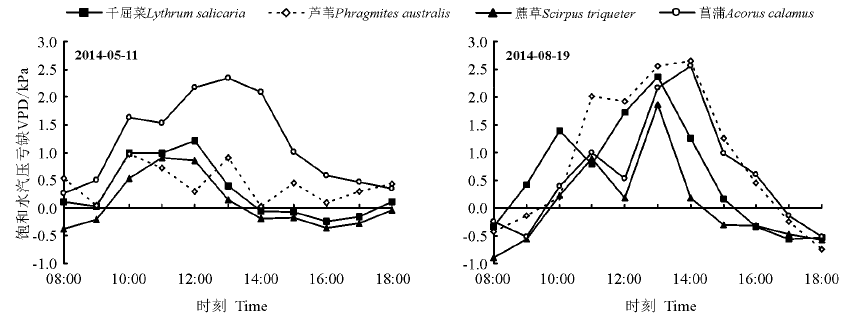 | 图6 4种植物水汽压差的日变化Fig.6 Diurnal saturation vapour pressure difference of 4 wetland plants |
5月11日千屈菜、芦苇的WUE呈双峰曲线, 均在12:00达到第一峰值(2.933 μ mol CO2· mmol-1 H2O、4.194 μ mol CO2· mmol-1 H2O), 在15:00达到第二峰值(2.775 μ mol CO2· mmol-1 H2O、3.488 μ mol CO2· mmol-1 H2O), 藨草、菖蒲的WUE波动变化, 没有明显的峰值。8月19日千屈菜、芦苇、藨草的WUE呈双峰曲线, 均在11:00时达到第一峰值(2.019、1.691和2.864 μ mol CO2· mmol-1 H2O), 在16:00时达到第二峰值(4.867、1.727和2.421 μ mol CO2· mmol-1 H2O), 菖蒲的WUE呈单峰曲线, 13:00达到峰值(4.467 μ mol CO2· mmol-1 H2O)。5月11日日均WUE表现为芦苇> 千屈菜> 藨草> 菖蒲, 8月19日表现为菖蒲> 藨草> 千屈菜> 芦苇(图7)。
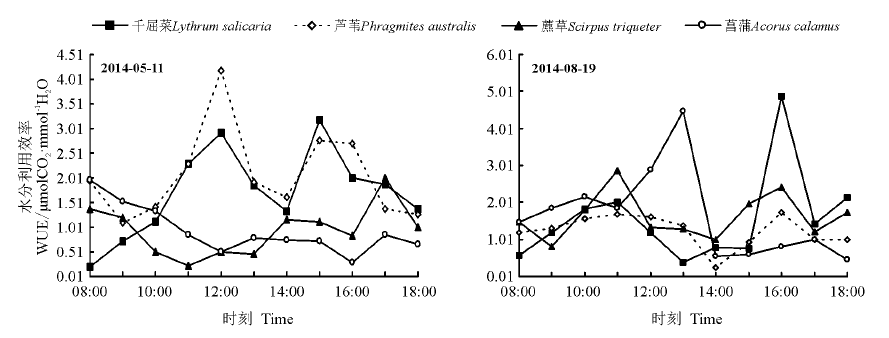 | 图7 4种植物水分利用效率的日变化Fig.7 Diurnal water use efficiency of 4 wetland plants |
对4种湿地植物的Pn与各生理因子和环境因子间进行相关性分析。结果表明, 5月11日千屈菜的Pn与PAR、Ta呈正相关(P< 0.1), 相关系数分别为0.619、0.509; 芦苇的Pn与PAR呈极显著正相关(P< 0.01), 相关系数为0.736; 藨草Pn与RH、PAR、Ta均呈正相关(P< 0.1), 相关系数分别是0.554、0.518、0.510; 菖蒲的Pn与Ca、RH呈显著正相关(P< 0.05), 相关系数分别为0.637、0.639(表1)。8月19日千屈菜、芦苇的Pn与PAR、Ta的相关性均不显著, 藨草的Pn与PAR呈极显著正相关(P< 0.001), 与Ca呈显著正相关(P< 0.05), 与SM呈极显著正相关(P< 0.01); 菖蒲的Pn与PAR、Ca呈显著正相关(P< 0.05), 与SM呈正相关(P< 0.1)。
| 表1 Pn与环境因子的相关系数 Table 1 Correlation coefficients between Pn and physiological factors |
植物Pn与生理因子间相关性分析显示, 5月11日千屈菜的Pn与Tr呈正相关(P< 0.1), 与Gs呈显著正相关(P< 0.05), 与WUE呈极显著正相关(P< 0.01), 相关系数分别为0.541、0.638、0.749, 与Ci 、VPD呈负相关; 芦苇的Pn与Tr呈显著正相关, 与Gs呈极显著正相关, 与WUE呈极显著正相关(P< 0.001), 关系数分别为0.622、0.720、0.821; 藨草的Pn与WUE呈极显著正相关, 相关系数为0.832; 菖蒲的Pn与Ci、WUE呈正相关, 相关系数分0.536、0.664。4种湿地植物的Pn与WUE的相关系数都较高。8月19日千屈菜的Pn与Tr呈极显著正相关, 与Gs呈正相关, 相关系数分别为0.715、0.529; 芦苇的Pn与各因子的相关性较小; 藨草的Pn与Tr呈极显著正相关(P< 0.001)、与VPD呈极显著正相关, 相关系数分别为0.879、0.752; 菖蒲的Pn与WUE呈极显著正相关(P< 0.001), 相关系数为0.921(表2)。
| 表2 Pn与生理因子的相关系数 Table 2 Correlation coefficients between Pn and physiological factors |
通径分析可以通过分解自变量与因变量间表面直接的相关性, 来研究自变量对因变量的直接影响和间接影响 [19], 对4种湿地植物Pn与环境和生理因子间相关性进行通径分析。
5月11日环境因子结果表明(表3):1)对千屈菜Pn影响的总效应表现为PAR> Ta> Ts> RH> Ca> SM, 即PAR对千屈菜Pn的影响最大(通径系数为0.595), 其中直接影响效应占主导, 为0.829, 对千屈菜Pn的直接效应表现为Ts> RH> Ca> PAR> Ta> SM; 2)对芦苇Pn影响的总效应表现为Ca> RH> PAR> Ta> SM> Ts, 即Ca对芦苇Pn的影响最大(通径系数为1.965), 其中直接影响效应占主导, 为-0.797, 对芦苇Pn的直接效应表现为PAR> Ts> Ta> SM> Ca> RH; 3)对藨草Pn影响的总效应表现为Ta> Ts> RH> Ca> PAR> SM, 即Ta对藨草Pn的影响最大(通径系数为-0.644), 对藨草Pn的直接效应表现为PAR> Ca> Ts> Ta> RH> SM, 即PAR对藨草的直接效应大于Ta的直接效应, 二者均通过彼此对Pn的间接影响共同构成对Pn的总效应[20]; 4)对菖蒲Pn的影响总效应表现为RH> Ca> Ts> SM> PAR> Ta, 即RH对菖蒲Pn的影响最大(通径系数为0.613), 其中直接影响效应占主导, 为1.422, 对菖蒲Pn的直接效应表现为Ts> RH> Ca> Ta> PAR> SM。每种植物的环境决定因子都存在差异性。
| 表3 Pn与环境因子的通径分析 Table 3 Path analysis between Pn and environmental factors |
8月19日环境因子结果表明(表3):1)对千屈菜Pn影响的总效应表现为PAR> Ca> SM> Ts> RH> Ta, 即PAR对千屈菜Pn的影响最大(通径系数为0.441), 其次是通过Ca和SM的间接效应; 对千屈菜Pn的直接效应表现为Ts> RH> Ta> SM> Ca> PAR; 2)对芦苇Pn影响的总效应表现为PAR> SM> Ta> Ca> RH> Ts, 即PAR对芦苇Pn的影响最大(通径系数为0.497), 对芦苇Pn的直接效应表现为RH> Ts> Ta> Ca> SM> PAR, RH对芦苇的直接效应大于Ts、Ta的直接效应, 它们通过彼此对对Pn的间接影响共同构成对Pn的总效应; 3)对藨草Pn影响的总效应表现为Ca> PAR> SM> Ta> RH> Ts, 即Ca对藨草Pn的影响最大(通径系数为1.432), 其中直接影响效应占主导, 为0.992, 其次是通过PAR和Ta的间接效应; 对藨草Pn的直接效应表现为Ca> RH> Ts> SM> PAR> Ta; 4)对菖蒲Pn影响的总效应为PAR> Ca> SM> RH> Ts> Ta, 即PAR对菖蒲Pn的影响最大(通径系数为0.637), 其中直接影响效应占主导, 为1.453, 其次是通过Ca和SM的间接效应; 对菖蒲Pn的直接效应表现为RH> Ts> Ca> Ta> PAR> SM, 即RH对菖蒲的直接效应大于PAR的直接效应, 二者同对Pn的间接影响构成对Pn的总效应。
5月11日生理因子结果表明(表4):1)对千屈菜Pn影响的总效应表现为WUE> Gs> Ci> VPD> Tr, 即WUE对千屈菜Pn的影响最大(通径系数为0.774), 其中直接影响效应占主导, 为0.736, 对千屈菜Pn的直接效应表现为WUE> Tr> Gs> Ci> VPD; 2)对芦苇Pn影响的总效应表现为WUE> Gs> Tr> VPD> Ci, 即WUE对千屈菜Pn的影响最大(通径系数为0.821), 其中直接影响效应占主导, 为0.798, 对芦苇Pn的直接效应表现为WUE> Tr> VPD> Gs> Ci; 3)对藨草Pn影响的总效应表现为WUE> VPD> Ci> Gs> Tr, 即WUE对藨草Pn的影响最大(通径系数为0.628), 其中直接影响效应占主导, 为1.088, 对藨草Pn的直接效应表现为WUE> Tr> VPD> Gs> Ci; 4)对菖蒲Pn影响的总效应表现为WUE> Gs> VPD> Tr> Ci, 即WUE对菖蒲Pn的影响最大(通径系数为0.246), 其中直接影响效应占主导, 为1.245, 对菖蒲Pn的直接效应表现为WUE> Tr> Gs> VPD> Ci。8月19日生理因子结果表明(表4):1)对千屈菜Pn影响的总效应表现为Tr> Gs> WUE> VPD> Ci, 即Tr对千屈菜Pn的影响最大(通径系数为0.704), 其中直接影响效应占主导, 为1.071, 对千屈菜Pn的直接效应表现为Tr> WUE> VPD> Ci> Gs; 2)对芦苇Pn影响的总效应为WUE> VPD> Tr> Gs> Ci, 即WUE对芦苇Pn的影响最大(通径系数为0.458), 对芦苇Pn的直接效应表现为WUE> Gs> Tr> Ci> VPD; 3)对藨草Pn影响的总效应表现为Tr> VPD> WUE> Gs> Ci, 即Tr对藨草Pn的影响最大(通径系数为0.866), 其中直接影响效应占主导, 为1.001, 对藨草Pn的直接效应表现为Tr> WUE> Gs> VPD> Ci; 4)对菖蒲Pn影响的总效应表现为WUE> Tr> Gs> VPD> Ci, 即WUE对菖蒲Pn的影响最大(通径系数为0.918), 其中直接影响效应占主导, 为0.934, 对菖蒲Pn的直接效应表现为WUE> Tr> Ci> VPD> Gs。
| 表4 Pn与生理因子的通径分析 Table 4 Path analysis between Pn and physiological factors |
通径分析是通过分析变量间的相关结构来定量地研究、解释变量间关系的一种方法[21], 具有相关分析和多元回归分析所不具备的优势[22]。与典型相关分析相比, 通径分析能够提供更多信息, 更客观, 更全面的揭示Pn与各影响因子的密切程度, 且能指出这种关系中哪种作用处于主导地位[23]。因此, 通径分析能全面反映自变量对因变量的相对重要程度[23, 24]。
本研究中, 5月11日千屈菜、芦苇的Pn是双峰曲线, 没有发生严重的“ 午休” 现象, 藨草和菖蒲的Pn变化较为平稳。8月19日千屈菜、芦苇、藨草的Pn是双峰曲线, 光合“ 午休” 现象明显, 这与大多数植物光合日变化趋势相似[25, 26], 菖蒲为单峰曲线。可能主要是由于5月11日的PAR明显低于8月19日, 而且午间光能对植物形成的光抑制也较小。同化高峰出现在上午, 与光照和气孔导度日变化规律相似, 这说明4种植物具有在不利环境条件出现之前, 可以在短暂时间内充分利用水和光源的能力, 这是对环境适应的表现[27]。
5月11日Pn由高到低依次是芦苇、千屈菜、菖蒲、藨草, 芦苇的最大Pn和Tr均较高。8月19日Pn由高到低依次是芦苇、菖蒲、千屈菜、藨草, 芦苇的最大Pn、Tr和Gs也均较高, 表明4种湿地植物中芦苇具有较强的光合能力, 段晓男等[28]在对乌梁素海野生芦苇光合特性的研究中发现, 芦苇的光合速率和蒸腾速率都高于附近的草原植物, 可以说明芦苇是高光合、高蒸腾的禾本科植物。5月11日菖蒲的Gs最高, 表明通过保持较高的气孔导度来保证较高的碳同化速率[20], 以适应湿地的气候条件。
环境因子中, 对千屈菜5月11日和8月19日的Pn影响最大的均为PAR, 它作为叶片能量的主要来源, 决定了一些环境与生理因子的变化。对芦苇5月11日和8月19日的Pn影响最大的分别为Ca和PAR。这是因为芦苇具有较强的光合能力, 5月份正是芦苇大量吸收CO2进行同化作用的时期。8月份属于生长季晚期, Ca对芦苇Pn的影响减弱而PAR的影响增强。对藨草5月11日和8月19日的Pn影响最大的分别为RH和Ca。这是由于PAR引起Ta变化进而影响RH, 导致叶内外蒸汽压差的变化, 对生长旺季的藨草正影响很大。8月份藨草正处于开花结实时期, 需要吸收CO2来积累碳水化合物。对菖蒲5月11日和8月19日的Pn影响最大的分别为RH和PAR。光合作用日变化曲线在不同物种之间存在着差异, 植株年龄、生育期、季节、环境条件、施肥状况也对峰型产生一定的影响[7, 29], 所以环境因子与光合作用间的关系并不是恒定不变的。
生理因子中, 对千屈菜、芦苇、藨草、菖蒲5月11日的Pn影响最大的均为WUE, 直接正效应占主导地位, 是4种植物光合作用的主要生理决定因子, 其次是Tr; 植物叶片水分利用效率作为植物生理活动过程中消耗水形成有机质的基本效率, 成为确定植物体生长发育所需要的最佳水分供应的重要指标之一[30]。有研究认为, 在干旱和半干旱地区WUE较高时表明植物能够成功或良好地生长[31], 这种自身水分调节功能使其既充分利用了当地丰富的光照资源, 又避免了因蒸腾过度使土壤更加干旱。表明4种湿地植物对当地的生态气候有很好的适应性[32]。对千屈菜和藨草8月19日Pn影响最大的是Tr, 直接正效应占主导地位; 对芦苇和菖蒲8月19日Pn影响最大的是WUE, 直接正效应占主导地位, 其次为Gs或Tr, 这与李林峰[5]得出的影响菖蒲和千屈菜Pn的主要因子为Gs和Tr相似, 本研究引入因子WUE, 在水资源缺乏的半干旱地区是一个重要的生理特征[31]。
环境和植物自身因素都会对植物光合生理特性产生影响[16]。通过相关性分析可知, PAR是最重要的影响因子。PAR影响Ta, RH、叶片温度又作用于Tr。在植物和环境相互作用的实际生态过程中, 依据物种和环境的变化, 各环境因子对植物产生的影响不是单一孤立和一成不变的, 而是互为制约, 综合作用[33]。
The authors have declared that no competing interests exist.
| [1] |
|
| [2] |
|
| [3] |
|
| [4] |
|
| [5] |
|
| [6] |
|
| [7] |
|
| [8] |
|
| [9] |
|
| [10] |
|
| [11] |
|
| [12] |
|
| [13] |
|
| [14] |
|
| [15] |
|
| [16] |
|
| [17] |
|
| [18] |
|
| [19] |
|
| [20] |
|
| [21] |
|
| [22] |
|
| [23] |
|
| [24] |
|
| [25] |
|
| [26] |
|
| [27] |
|
| [28] |
|
| [29] |
|
| [30] |
|
| [31] |
|
| [32] |
|
| [33] |
|