
{kind=link}
{kind=link}
{kind=link}
{kind=link}
高寒草甸植被特征对模拟增温的响应——以青藏高原多年冻土区为例
[陈翔1, 2  , 彭飞
, 彭飞1 , 尤全刚1 , 王涛1 , 薛娴1 ]
, 彭飞, 尤全刚, 薛娴|
|


第一作者:陈翔(1991-),女,甘肃陇西人,在读硕士生,主要从事全球变化生态研究。E-mail: [email protected]
采用红外辐射灯(infrared heater)模拟气候变暖背景,研究青藏高原多年冻土区高寒草甸优势植物种珠芽蓼( Polygonum viviparum)、美丽风毛菊( Saussurea superb)和黑褐苔草( Carex atrofusca)的生长发育及光合特性对气候变暖的短期响应及其差异性,旨在为评价草地生态系统的敏感性和脆弱性提供科学依据。结果表明,模拟增温W1(1.88 ℃)和W2(3.19 ℃)均可改变高寒草甸物种分布的频度和季节分布格局,并显著影响植被高度和光合色素含量,影响程度存在种间差异性。与不增温对照相比,W1增温下物种具有不同的响应,美丽风毛菊的高度、频度和叶绿素含量均呈下降趋势,而珠芽蓼和黑褐苔草均呈增加趋势。W2增温显著增加了珠芽廖和黑褐苔草的高度、频度和叶绿素含量( P<0.05),却显著降低了美丽风毛菊的叶绿素a/b值( P<0.05)。物种间光合特性存在显著差异( P<0.05),样地间差异性表现更为突出,但增温处理间变化不明显。这说明增温可促进高寒草甸牧草类植物的生长发育,延长青草期,有利于牧业生产,但斑块状分布明显。
Infrared heater was used to simulate temperature enhancement in the field focusing on the effects of temperature enhancement on the growth and photosynthetic characteristics of three alpine plants: Polygonum viviparum, Saussurea superb, and Carex atrofusca in order to provide basis for estimating the sensitivity and friability of the meadow ecosystem in the permafrost region on the Qinghai-Tibet Plateau, China. The results showed that the simulated warming W1 (1.88 ℃) and W2 (3.19 ℃) could all alter the distributing frequency and season pattern of the alpine plant species. Compared with the CK, the W1 temperature enhancement brought different changes to different plants under which the height, frequency and chlorophyll contents of the S. pulchra were decreased but the values of the P. viviparum and C. atrofusca were increased. The W2 warming evidently increased the height, frequency, and chlorophyll content of the P. viviparum and C. atrofusca but significantly ( P<0.05) decreased the ratio of chlorophyll a to b of the S. pulchra resulted in its decreases in the height, frequency, and chlorophyll content. It showed significant differences in the photosynthesis indicators among the plant species, especially among the plots. All above indicates that the warming can improve the growth and development of the grazing plants in the alpine meadow and prolong their green grass duration resulting in benefit to the livestock production but the vegetation distribution evidently variegated.
自1850年以来, 全球表面温度平均升高了0.74 ℃, 并且据模型估算到21世纪末全球表面平均温度将升高1.8~4.0 ℃[1, 2]。碳耦合模型以及气候观测资料均表明青藏高原升温幅度大于全球平均值[1, 2, 3]。与其它地区冻土相比, 青藏高原冻土纬度低、海拔高、接受辐射强、冻土温度高, 是全球气候变化响应的敏感区和脆弱区, 极易受到气候变暖和人类活动的影响[4]。
温度控制着生态系统中许多生物化学反应过程的速率, 影响着几乎所有生物学过程。低温和短的生长季是高寒生态系统生产力主要限制因子。因此, 直接升高温度和间接的生长季延长在很大程度上可能促进高山植物的生长发育, 进而改变群落的组成和结构[5]。增温对植物的影响表现为直接影响和间接影响。增温可直接改变植物的光合作用能力和生长速率[6], 从而改变植物的物候[7]和节律, 并延长植物的生长期[8]。间接的影响包括增温后土壤理化特性的变化对植物生长、生物量生产分配的影响[9, 10]和土壤微生物群落组成及多样性的改变对植物产能的影响。增温对植物的直接或间接影响都会改变高寒生态系统的碳循环。高寒地区冻土碳库是全球重要的陆地生态系统碳库之一。据估算, 高寒地区冻土储存了约1 672 Pg碳(1 Pg=1 015 g)[11, 12], 其中的12%是近地表活动层土壤有机碳, 88%是冻土碳库[12]。冻土有机碳分解是高寒生态系统土壤碳通量的一个重要组成部分。冻土生态系统光合固定碳和呼吸释放碳之间的平衡影响着气候变化的强度和方向[13]。
草甸植物生长特性和光合特性是碳循环研究的重要内容, 已经成为植物生理学家与生理生态学家共同关注的课题[14]。红外辐射(infrared radiation)技术模拟增温可以同时改变显热、潜热和土壤热通量, 且对土壤物理干扰小, 是理想的模拟气候变暖研究方法[15]。模拟气候变暖研究涉及面较广。钟永德等[16]研究指出, 地球暖化促进了植物的迁徙与入侵。然而, 目前模拟增温研究[14, 15, 16, 17, 18, 19, 20, 21, 22]大都是进行增温与对照的单一比较研究, 而对多个梯度水平温度的定量影响研究相对不足。对青藏高原高寒草甸光合生理研究多基于叶片水平, 而群落水平的研究也相对匮乏[14]。也有学者[3, 23, 24]采用红外辐射增温方法研究了青藏高原土壤温度和碳循环的变化, 发现增温处理使土壤表层含水量降低, 却使耕层下土壤含水量增加, 指出模拟增温造成植物生存的土壤环境发生改变, 但利用红外辐射增温装置模拟不同增温梯度下草甸植物群落的分布与生长特性的研究尚少见报道。因此, 从群落尺度研究青藏高原典型高寒草甸植被物种在不同模拟增温条件下的变化规律及其驱动机制均具有重要意义, 可为评价草地生态系统的敏感性和脆弱性提供科学依据。
研究区位于中国科学院冻土工程国家重点实验室青藏高原研究基地(简称北麓河试验站), 地理坐标为34° 49'33.98″-34° 49'36.58″ N, 92° 55'57.19″-92° 56'06.32″ E, 平均海拔4 630 m。该地属青藏高原干旱气候区, 年均气温为-5.9 ℃, 年均降水量267.6 mm, 年均潜在蒸散量1 316.9 mm, 年均风速4.1 m· s-1, 冻结期为9月至翌年4月。植被属高寒草甸类型, 以高山嵩草(Kobresia tibetica)为建群种, 伴生有矮火绒草(Leontopodium nanum)、美丽风毛菊(Saussurea pulchra)、藏嵩草(K. tibetica)、黑褐苔草(Carex moorcroftii)、细小棘豆(Oxytropis pusilla)等, 以莎草科和菊科的植物为主, 蓼科的珠芽蓼(Polygonum viviparum)也较为常见, 植被盖度达83%[20]。
模拟增温试验采用红外辐射法[3, 23]进行, 即对样地的增温通过安装红外辐射器(MR-2420, Kalglo Electronics Inc., Bethlehem, PA, USA)来实现。试验采用随机区组设计, 5次重复, 即沿海拔梯度划定5个区组, 同一区组内海拔高度基本一致, 并尽可能选择样地的微地形、土壤状况和植被组成上的高度相似, 以提高模拟增温试验的精确性和区组样地的代表性。区组Ⅰ 和区组Ⅱ 位于等海拔区域内, 海拔高度变幅为4 633.5~4 634.0 m, 区组Ⅲ 和Ⅴ 位于等海拔区域内, 海拔变幅4 633.0~4 633.5 m。区组间相距50 m左右。每个区组内随机布设W1(1.88 ℃)和W2(3.19 ℃)2个增温样方和1个对照样方(CK), 样方面积2 m× 2 m, 每个增温样方内距地表1.5 m处悬挂1个红外辐射器(165 cm× 15 cm)向下辐射增温, 对照不增温样方内悬挂同形不锈钢环以消除增温器误差。每个区组内除增温处理外, 其它环境条件基本一致。以样地内的主要植物种美丽风毛菊(Saussurea pulchra)、黑褐苔草(Carex moorcroftii)和珠芽蓼(Polygonum viviparum)为研究对象于生长季测量增温对这3种植被的生理和生态指标的影响。
1.3.1 物种频度和高度测定 在植物生长盛期7-9月每月上旬在试验样方内测定一次物种的频度和高度。植被高度用特制的直尺测定, 由于高寒草甸植物矮小, 为减小测量误差, 将直尺零刻度线至边缘的空白区去掉, 使直尺从零刻度开始, 每一个样方内进行测量时, 在样方两条交叉对角线测定高度值, 每条对角线上随机测定20株, 共测定40株高度值, 以平均值
采用一个27 cm× 27 cm的样方框测定频度。样方框内有100个2.5 cm× 2.5 cm的小网格。测定时将样方框平铺到各样方中央地面, 然后统计每个样方内每个物种出现的小方格数, 计为频度(%)。如果样方框内没有该物种, 频度数计为0。最后计算每种处理下5个重复的平均数
CV=SD/
1.3.2 光合速率测定 由于研究区植物生长季开始晚, 生长期短, 5月开始返青, 11月开始萎黄越冬, 植物个体通常在7-8月份光合生理指标相对稳定[25]。 在2011年7月和8月及2012年7月份选择晴朗的天气测定2~3次光合生理指标, 共测定9次, 9次测量的平均值作为该植物的光合生理参数。光合生理指标采用便携式光合仪(Li-6400 xt, Li-cor, Inc.Lincoln, NE, USA)测定, 测定方法及过程如下:
在各区组样地对照样方和增温样方中, 分别随机选取3种植物样株各3株, 用便携式光合仪在晴天的上午或者下午光照充足且相对稳定的时间内测定完全展开的叶片的光合作用, 每株重复测定5次。珠芽蓼单个叶片可以填充满叶室, 直接测定, 美丽风毛菊和黑褐苔草两个物种的叶片面积都小于6 cm2, 这两个物种的叶片长度大于3 cm, 测定时在叶片的根部用胶带将叶片并排粘在一起以填充满叶室, 然后扫描依次读取3种植物的瞬时净光合速率、蒸腾速率、气孔导度、胞间CO2浓度等光合特性指标。
1.3.3 叶绿素含量测量 将2011年7月份用于测定光合作用的叶片放入2 mL的离心管中。所有的离心管放入保鲜盒带回实验室。取不同物种的植物叶片, 擦干净叶片表面的污物, 用二次蒸馏水冲洗, 剪碎(去掉中脉)。称取每种植物的新鲜样品0.2 g, 共3份, 分别放入玛瑙研钵中, 加入少量的石英砂及2~3 mL 95%乙醇, 研成浆, 再加乙醇10 mL, 继续研磨至叶片组织变白, 静置3~5 min。取一张滤纸, 置漏斗中, 用乙醇湿润, 沿玻璃棒把提取液导入漏斗中, 过滤到25 mL棕色容量瓶中, 用少量乙醇冲洗研钵, 研棒及残渣数次。用移液枪吸取乙醇, 将滤纸上的叶绿体色素全部冲洗如容量瓶中。最后用乙醇定容至25 mL, 摇匀。把叶绿素提取液倒入石英比色杯中。以95%乙醇为空白, 在波长665和649 nm下测定吸光度[26]。
叶绿素a=[137× (A665-0.055 011)-5.76× (A649-0.053 367)]/鲜叶重;
叶绿素b=[25.8× (A649-0.053 367)-7.6× (A665-0.055 011)]/鲜叶重;
叶绿素=叶绿素a+叶绿素b。
式中, A665和A649为叶绿素溶液在波长665 nm和波长649 nm时的吸光度。
增温对植被生理和生态参数的影响采用双因素随机区组试验设计的方差分析。将物种和增温处理分别作为试验因子, 研究物种间和处理间的差异, 同时研究这两个因素的交互效应来揭示增温在不同物种间是否存在差异。方差分析采用SPSS 17.0软件进行, 多重比较选用Duncan法。文中图的绘制采用Excel 2003版软件完成, 采用标准误作误差线。
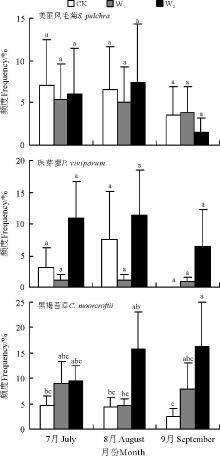
物种频度是反映植被群落特征的重要指标。在本研究各区组样地15个样方范围内, 平均而言, 物种频度从高到低依次为黑褐苔草(8.29%)> 美丽风毛菊(4.91%)> 珠芽蓼(4.76%)。W2处理下3种草甸物种平均频度均最高 (9.47%, n=15, CV=129.4%), 对照居中(4.38%, n=15, CV=111.6%), W1处理下3种物种平均频度均最低(4.11%, n=15, CV=149.3%)。增温处理对3种物种出现频度的影响也因物种的不同而异(图1), 随模拟增温程度加大黑褐苔草频度呈持续增加趋势, W1和W2使黑褐苔草频度分别较对照提高了89.5%和264.9%, 美丽风毛菊和珠芽蓼频度基本呈先下降后增加的趋势, 珠芽蓼频度变异程度最大(CV=184.7%), W1较对照下降68.5%, W2较对照提高164.8%。对照条件下, 随着生长季节的延后, 高寒草甸优势种美丽风毛菊和黑褐苔草频度均呈下降趋势, 珠芽廖在8月频度最高(图2)。W1增温降低了美丽风毛菊和珠芽蓼的频度, 但差异未达到显著水平(P> 0.05), W2增温降低了美丽风毛菊的频度, 却增加了黑褐苔草和珠芽蓼的频度, 其中对黑褐苔草频度的正效应达显著水平(P< 0.05), 且随生长季节的延后趋势愈加明显(图1, 2)。
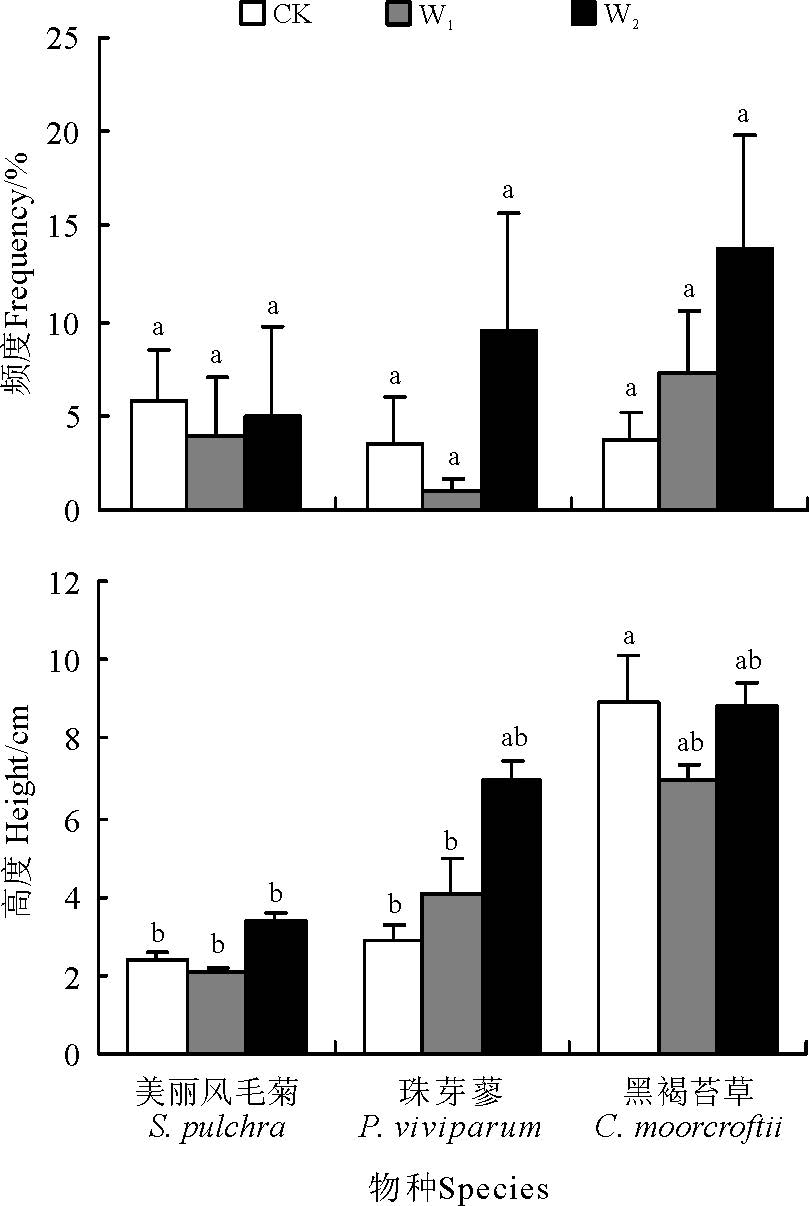 | 图1 植被高度和频度对模拟增温响应的比较 注:W1, 增温1.88; W2, 增温3.19 ℃。不同小写字母表示不同物种与不同增温处理组合间差异极显著(P< 0.05)。下同。Fig.1 Response comparison of vegetation height and frequency to the simulated warming Note: W1 and W2 simulated warming 1.88 and 3.19 ℃, respectively. Different lower case letters show great significant difference among combinations of different species and warming treatments at 0.05 level. The same below. |
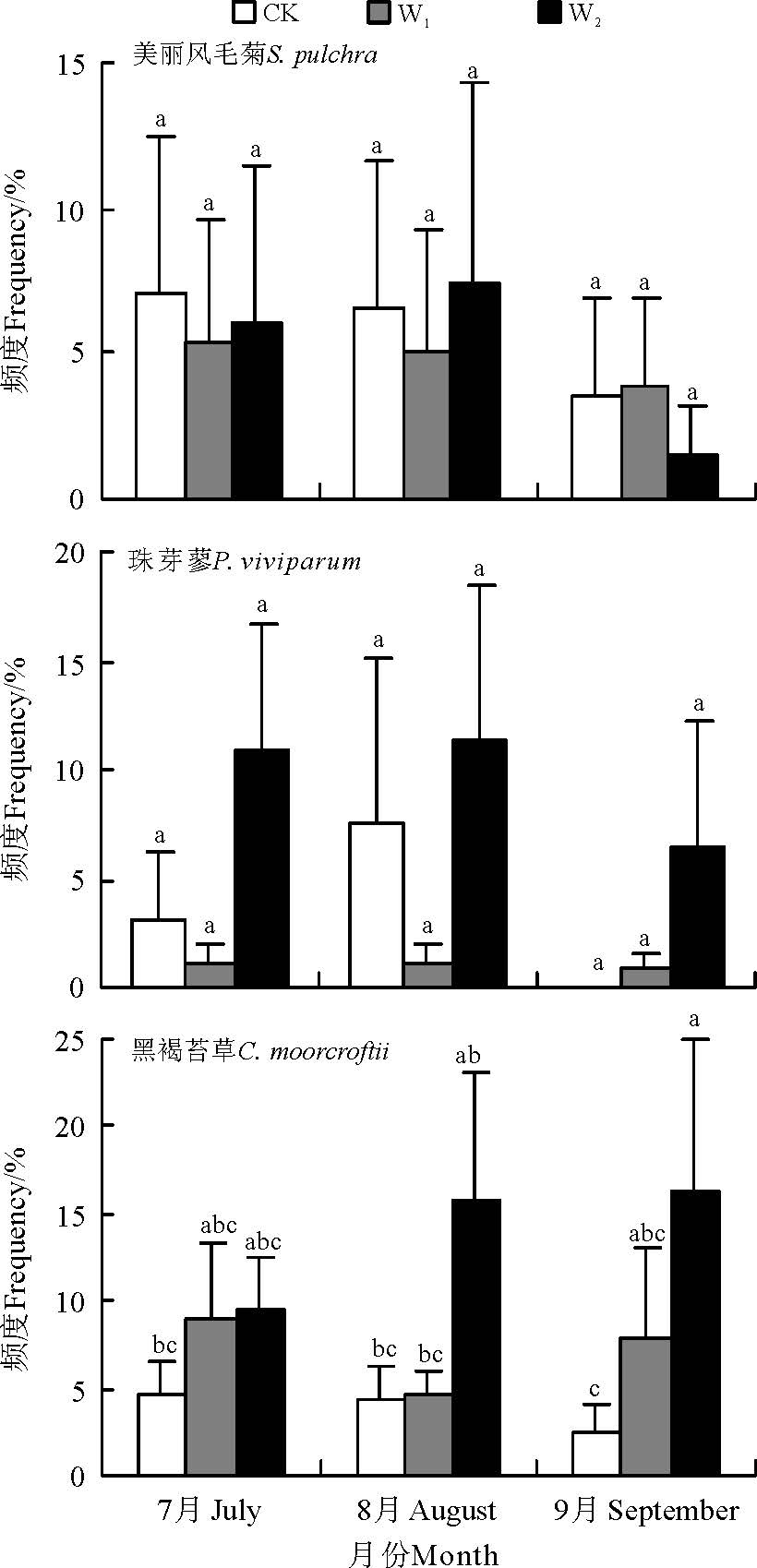 | 图2 高寒草甸3种植物频度对模拟增温的响应Fig.2 Response of vegetation height and frequency to the simulated warming |
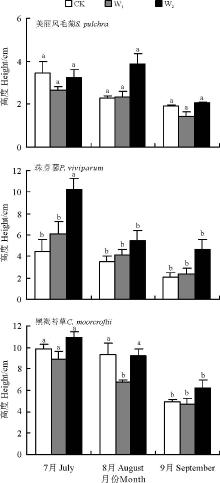
高寒草甸3个物种间植被平均高度存在显著差异(F=41.542, P< 0.01), 植株高度从高到低依次为黑褐苔草(8.31± 0.39, n=14, CV=22.37%)> 珠芽蓼(4.87± 0.46, n=10, CV=45.61%)> 美丽风毛菊(2.51± 0.52, n=8, CV=24.06%)。W2处理下3种草甸植被平均高度最高(7.13± 2.31, n=11, CV=32.42%, 95%置信区间为5.37~7.32 cm), 对照处理的水平居中(5.53± 3.68, n=11, CV=66.49%, 95%置信区间为3.84~5.69 cm), W1处理下植被平均高度最低(4.61± 2.41, n=10, CV=52.36%, 95%置信区间为3.40~5.40 cm)。增温处理对3种物种植被高度的影响也因物种的不同而异, 珠芽蓼对增温最为敏感, 而美丽风毛菊和黑褐苔草植被高度先降低后增加, 响应机制较为一致(图1, 3)。在7-9月, 各增温处理中, 物种植被高度随生长季节的延后均呈减小趋势(P=0.05), 7月植株最高, 8月开始减小, 9月最低。降低程度也因物种的不同而异, 其中, 美丽风毛菊下降程度均未达显著水平(P> 0.05), 黑褐苔草的降低程度均达显著水平(P< 0.05), 珠芽蓼在W2增温样地植被高度显著下降, 而在W1和不增温样地, 下降程度不显著。与对照相比较, 随增温加大黑褐苔草的频度和珠芽廖的高度呈持续增加趋势, W2使珠芽廖频度明显增加, 高度显著提高。
对物种频度和高度的方差分析表明, 试验区组间物种频度与高度差异均不显著(P> 0.05)。3个物种间与3个增温处理间频度的差异性也均未达到显著水平(表1), 而3个增温处理间物种高度的差异达到极显著水平(P< 0.01)。物种与增温处理的互作效应对频度与高度的影响均不显著。
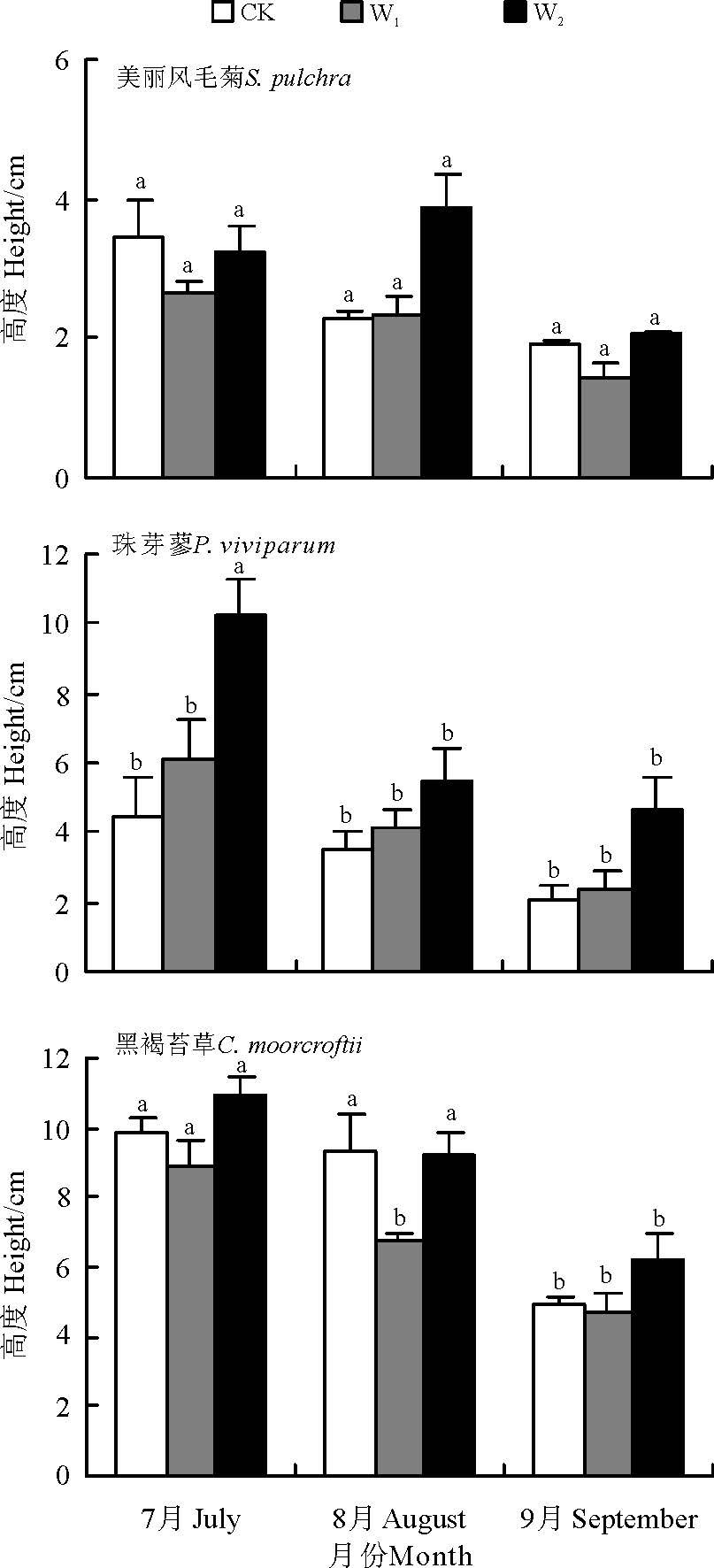 | 图3 高寒草甸3种植物高度对模拟增温的响应Fig.3 Responses of vegetation height to the simulated warming |
| 表1 模拟增温与物种间物种频度和植被高度的方差分析结果(F值) Table 1 Results (F value ) of the effects of warming and species on plant frequency and plant height |
增温处理对草甸植被叶绿素含量具有极显著影响(P< 0.01)(表2), 影响程度因物种和叶绿素种类的不同而异。草甸植被叶绿素含量从高到低依次为黑褐苔草> 珠芽蓼> 美丽风毛菊。W1增温显著提高了黑褐苔草的叶绿素a、叶绿素b和总叶绿素含量(P< 0.05), 显著降低了美丽风毛菊的叶绿素a含量。W2增温显著提高了美丽风毛菊a、b和总叶绿素含量及黑褐苔草的叶绿素b和总叶绿素含量, 增温处理对珠芽蓼的叶绿素含量影响不大(图4)。增温显著降低了叶绿素a/b值, 降低程度随物种的不同而异, 随模拟增温的升高, 珠芽蓼叶绿素a/b值先升高后降低, 而黑褐苔草和美丽风毛菊a/b值先下降后又升高, 美丽风毛菊变化更加显著(图4)。
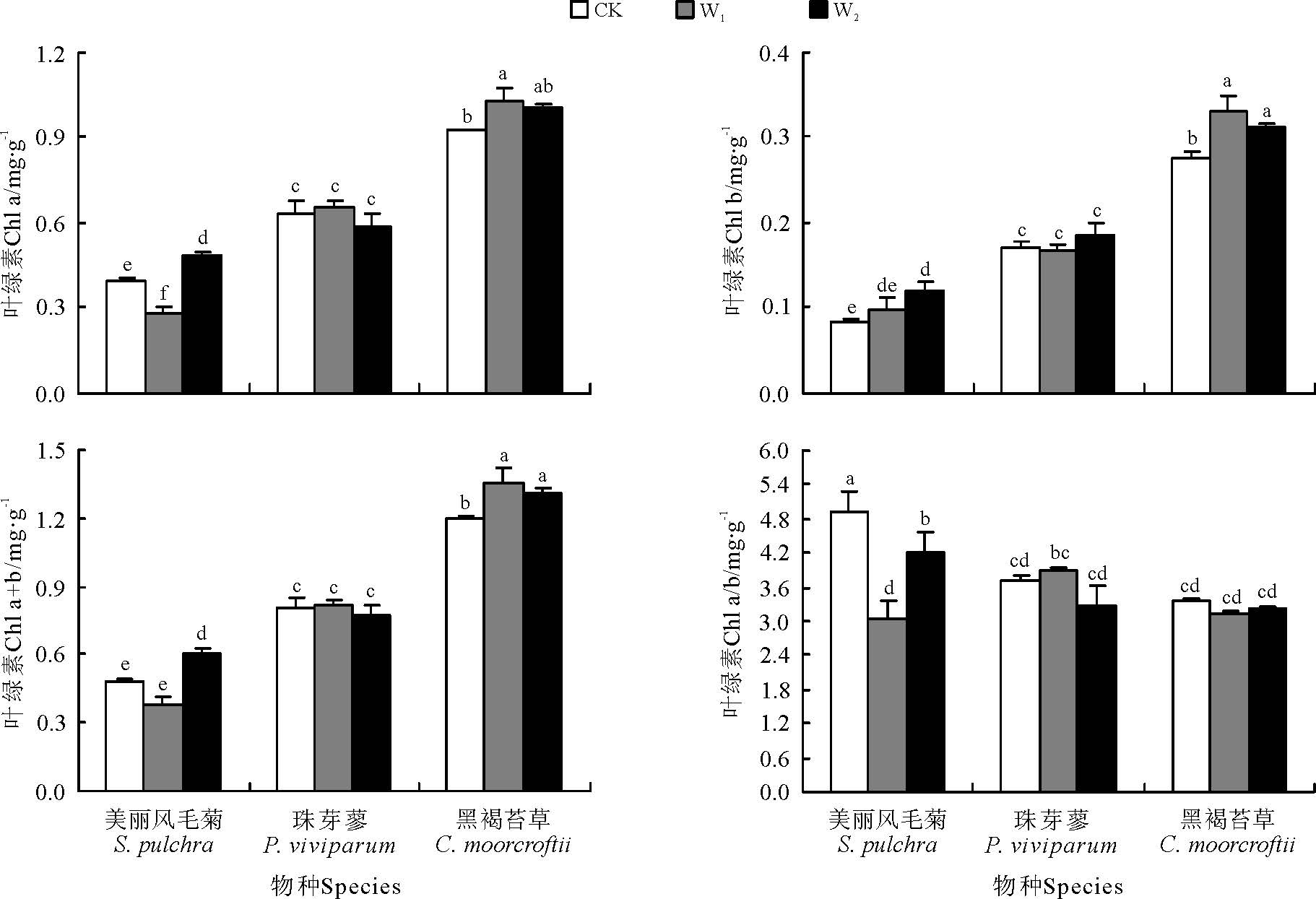 | 图4 不同草甸植物叶绿素对模拟增温的响应Fig.4 Responses of chlorophyll in different alpine meadow plants to the simulated warming |
方差分析结果表明, 叶绿素a和总叶绿素含量在区组间和物种间的差异性均不显著(P> 0.05), 而叶绿素b和a/b在物种间的差异性均达到显著水平(P< 0.05)。增温处理间物种叶绿素含量的差异性达到极显著水平(P< 0.01)。物种与增温处理对叶绿素a、总叶绿素、叶绿素a/b的互作效应也达到极显著水平(P< 0.01), 而对叶绿素b含量的互作效应则不显著(P> 0.05)(表2)。
| 表2 3个模拟增温处理与3种高寒草甸植物叶绿素含量的方差分析 Table 2 Variance analysis of the vegetation chlorophyll of three alpine plants under three simulated warming treatments |
在不同增温处理条件下, 美丽风毛菊和珠芽蓼的净光合速率、蒸腾速率、气孔导度和胞间CO2浓度虽出现差异, 但其差异性均未达到显著水平(P> 0.05)。黑褐苔草的净光合速率、蒸腾速率和气孔导度在3种增温处理下都显著小于美丽风毛菊和珠芽蓼的(P< 0.01), 只有胞间CO2浓度差异不显著(P> 0.05)(表3)。本研究范围内, 物种间光合特性存在极显著差异(P< 0.01), 增温对3种植物的净光合(P=0.643)和蒸腾速率(P=0.714)均没有显著影响(表5)。
光合特性指标间的相关分析(表4)表明, 不同增温处理条件下, 净光合速率与蒸腾速率和气孔导度均呈极显著正相关(P< 0.01), 而与胞间CO2浓度间的相关性不显著(P> 0.05)。在W1增温和对照条件下, 气孔导度与蒸腾速率和胞间CO2浓度间均呈极显著正相关, 增温条件下其相关程度均增大, 当增温温度升高至W2后出现差异, 气孔导度与胞间CO2浓度间呈显著正相关(P< 0.05), 但与蒸腾速率间的正相关达极显著水平(P< 0.01)。
对光合指标方差分析结果(表5)表明, 物种蒸腾速率在区组间的差异性达到显著水平(P< 0.05), 其余光合特性指标在区组间的差异性均达到极显著水平(P< 0.01)。3种物种间气孔导度、净光合速率、蒸腾速率的差异性均达到极显著水平(P< 0.01), 而胞间CO2浓度的差异性均不显著(P> 0.05)。3个增温处理间物种的光合特性指标差异性均不显著(P> 0.05), 物种与增温处理的互作效应也未达到显著水平, 说明高寒草甸物种的光合特性随模拟增温处理和物种的不同而异, 物种光合特性对模拟增温的响应机制趋于一致, 但影响的程度存在种间差异。
| 表3 增温对3种植物主要光合参数的影响 Table 3 Effects of main photosynthetic parameters to the simulated warming |
| 表4 不同模拟增温条件下3种高寒草甸植物主要光合指标间的相关性 Table 4 Correlations between the main photosynthetic indexes to the simulated warming |
| 表5 模拟增温与物种间植物主要光合指标的方差分析结果(F值) Table 5 Result (F value) of warming and species on main photosynthetic indicators of three alpine plants |
地球暖化可促进植物的迁徙与入侵[16]。位于青藏高原腹地的高寒草甸生态系统目前植物所处的环境温度普遍低于植物生长的最优温度, 因此增温通常都会促进植物的生长和生物量积累[27]。周华坤等[28]研究表明, 在温室内气温、地表温度、土壤层温度分别增加1.47、1.54和1.00 ℃的增温条件下, 组成祁连山南坡的高寒草甸主要种群的高度、盖度和重要值均有提高, 种群结构发生变化, 虽然杂草生物量减小了21.13%, 但是由于禾草和莎草地上生物量增加, 总地上生物量增加3.53%。而在川西地区的研究发现, 杂草牛尾蒿(Artemisia dubia)(菊科蒿属)的地上生物量显著增加[5]。李娜等[29]研究发现, 在整个生长季内较对照提高2.98 ℃时, 高寒草甸与沼泽草甸总生物量均增加, 但5.52 ℃时抑制了高寒草甸的这种促进作用。赵建中等[30]研究揭示, 高寒草甸的建群种植物的生物量随增温幅度的升高(2.1、2.2、2.9 ℃)先增大后减小。本研究中, 珠芽蓼、美丽风毛菊和黑褐苔草物种频度在样方间和季节间变异均较大, 强变异显示出3个物种群落结构的时空镶嵌性较突出, 造成群落在植被特征上表现为明显的斑块状分布。自然条件下, 7-9月随生长季的延后, 美丽风毛菊和黑褐苔草频度均呈下降趋势, 珠芽廖在8月频度最高。随增温的升高, 物种频度先减小后增大, 增温降低了美丽风毛菊的频度, 却增加了黑褐苔草和珠芽廖的频度, 且随生长季的延后愈加明显。增温对物种高度的影响极显著, 种间差异性更加明显, 珠芽蓼对增温最为敏感, 而美丽风毛菊和黑褐苔草高度响应机制较为一致。随增温强度加大, 黑褐苔草的频度和珠芽廖的高度呈持续递增趋势, W2使珠芽廖频度较对照迅速增加, 高度显著提高。说明模拟增温可改变高寒草甸物种分布频度和季节性分布格局, 并促进植物的生长, W2增温更有利于高寒草甸植物的生长发育, 这也印证了石福孙等[27]的研究结果, 这可能是因为增温促进了高寒草甸土壤中碳磷循环[31], 改善了土壤理化特性的缘故。
植物叶片的光合作用与呼吸作用的平衡决定着净碳固定, 对于气候变化与生态系统碳循环的反馈具有重要的意义。叶片尺度净光合增加是生物量增加的主要机制[3]。Liang等[32]研究表明, 增温可使叶片的净光合平均增加6.13%。C4植物对增温的正响应强度大于C3植物。环境温度越高, 增温对叶片净光合的正效应越低。不同物种的净光合对增温的影响存在差异, 禾草光合速率增温后增加了12.3%, 而杂草光合速率变化不明显。经过长期的演化适应, 高寒草甸植被一般细胞内叶绿体较小, 数量多, 使得光合膜面积增加, 有利于更多光合产物的合成[33]。本研究中, 模拟增温对草甸植被叶绿素含量具有极显著影响, 物种光合色素对增温的响应存在显著的互作效应, W1条件下黑褐苔草增加程度最大, 总叶绿素含量较对照提高13.15%, W2条件下美丽风毛菊增加程度最大, 总叶绿素含量较对照提高25.55%, 高寒草甸光合特性对模拟增温的响应机制存在显著的种间差异性和样地差异性, 但增温对3种植物的光合特性指标均无显著影响。各增温条件下黑褐苔草的净光合速率、蒸腾速率和气孔导度均最小, 美丽风毛菊和珠芽蓼接近。净光合速率与蒸腾速率、气孔导度间均呈极显著正相关, 而与胞间CO2浓度间的相关性不显著。说明在气候变暖背景下, 物种叶绿素含量提高, 叶绿素a/b值下降, 但光合特性指标变化不明显, 这可能是因为环境温度越高, 增温对高寒草甸植物叶片光合特性的正效应越低的缘故, 这与卢存福和贲桂英[33]的研究结果相一致。
综上所述, 美丽风毛菊、黑褐苔草和珠芽廖是构成青藏高原高寒草甸的优势植物种, 模拟增温可改变3种植被的频度分布。随着本研究中模拟温度的增加, 均促进了3种高寒草甸植被的生长, 在全球变暖背景下有限的增温范围内, 模拟增温延后了黑褐苔草和珠芽廖的生长季和青草期, 提高了生长量, 有利于畜牧业生产。然而本研究中仅有两个模拟增温梯度, 还需进一步考虑当温度持续增高时物种的响应。另外, 模拟增温处理后的样方其微气象特征均会发生变化, 本研究仅讨论了温度的变化情况, 更多的微气象因素包括光照、水分等有待进一步观测和分析, 下一步研究的重点将是对高寒草甸物种群落组成和土壤微生态系统的研究。
The authors have declared that no competing interests exist.
| [1] |
|
| [2] |
|
| [3] |
|
| [4] |
|
| [5] |
|
| [6] |
|
| [7] |
|
| [8] |
|
| [9] |
|
| [10] |
|
| [11] |
|
| [12] |
|
| [13] |
|
| [14] |
|
| [15] |
|
| [16] |
|
| [17] |
|
| [18] |
|
| [19] |
|
| [20] |
|
| [21] |
|
| [22] |
|
| [23] |
|
| [24] |
|
| [25] |
|
| [26] |
|
| [27] |
|
| [28] |
|
| [29] |
|
| [30] |
|
| [31] |
|
| [32] |
|
| [33] |
|