

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
围封对黄土高原草地土壤铵态氮和硝态氮的影响
[廖圣祥1 , 任运涛1 , 袁晓波1 , 李旭东1  , 郭丁
, 郭丁1 , 李金博2 , 傅华1 ]
, 郭丁|
|


第一作者:廖圣祥(1991-),男,安徽合肥人,在读硕士生,主要从事草地生态学研究。E-mail:[email protected]
本研究以黄土高原长期放牧草地和围封7年的草地为对象,分析了土壤铵态氮和硝态氮含量的季节变化,并应用RDA冗余分析法分析了土壤水分、植被地上绿色生物量以及土壤微生物生物量碳对土壤铵态氮、硝态氮的影响。结果表明,铵态氮是围封与放牧草地土壤有效氮的主要组分;两样地0-10 cm土层土壤铵态氮与硝态氮均主要受植被地上绿色生物量的影响;围封草地10-20 cm土层土壤硝态氮主要受土壤水分与植被地上绿色生物量的共同影响,而放牧草地10-20 cm土壤铵态氮和硝态氮主要受植被地上绿色生物量的影响;土壤铵态氮与硝态氮之间呈极显著正相关( P<0.01);放牧草地围封7年后,较高的生物量使得凋落物及土壤有机质增加;另外,围封后土壤水分在多数季节显著增加( P<0.05),这些因素共同促进了土壤氮素的矿化。
A study was conducted in fenced grassland (F, fenced for 7 years) and grazed grassland (G) on the Loess Plateau to investigate the seasonal variation of the contents of soil ammonium nitrogen(N-N) and nitrate nitrogen (N-N). Redundancy analysis was also used to study the effects of soil water content, above-ground green biomass and microbial biomass carbon on N-N and N-N contents. The results showed that N-N was the main component of soil available nitrogen in both fenced and grazed grasslands. In the 0-10 cm soil layers of the two sites, soil N-N and nitrate nitrogen were mainly controlled by above-ground green biomass. In the 10-20 cm soil layers, soil N-N and nitrate nitrogen were mainly controlled by both of soil water content and above-ground green biomass in fenced grassland, while soil N-N and N-N were mainly controlled by above-ground green biomass in grazed grassland. There was significantly positive correlations ( P<0.05) between N-N and N-N in the two sites. The results indicated that the higher above-ground biomass after fencing for 7 years increased litter inputs and soil organic matter, thus increased substrate for nitrogen mineralization. In addition, soil water content significantly increased in most seasons ( P<0.05) and all of these factors promoted the nitrogen mineralization in the soil.
氮是植物有机合成的重要营养元素[1, 2], 一方面它是植物从土壤中吸收最多的元素, 另一方面它又最容易通过淋溶或挥发而损失[3]。土壤中的氮素大部分以有机态存在(占全氮的85%~95%), 只有通过矿化转化为有效氮后才能被植物吸收和利用[4]。铵态氮和硝态氮是土壤有效氮的主要形态[5], 其被植物吸收的量约占阴、阳离子吸收总量的70%[6]。因此, 研究植物氮素营养时, 铵态氮和硝态氮便成了人们关注的焦点。
放牧是草地最主要的利用方式[7], 放牧活动会影响土壤中氮素的形态及转化过程, 但对其如何影响土壤氮素矿化还存在很大争议[8]。有研究表明, 轻度放牧会加快土壤氮矿化速率, 因为家畜的采食作用刺激了根系生长和分泌, 导致根际微生物活性增加, 从而导致土壤氮矿化速率升高[8]; 也有研究表明, 放牧家畜的践踏和采食作用会对植被地上活体部分、植被的功能群生物量以及植物群落的物种多样性产生一定的影响[9], 使得凋落物质量变差而降低土壤质量[10, 11], 进而影响土壤氮素的矿化。土壤肥沃地区其氮矿化潜力通常较高, 反之亦然[12]。当放牧强度超过一定阈值后, 植被生物量降低, 地上部分受损严重, 减少了根系分泌物, 从而导致土壤氮矿化速率下降[13, 14]。放牧活动可能主要是通过影响土壤水分和土壤温度而影响土壤氮素矿化[15]。
黄土高原是世界范围内土层厚度最大, 黄土分布面积最广的地区。长期以来, 由于人类不合理的利用和黄土的特殊性质, 使该区土壤侵蚀、植被退化和水土流失现象严重[16], 成为我国土壤氮素储量最低的地区之一[17]。围封是退化草地恢复的重要措施之一, 但是关于黄土高原围封与放牧对草地土壤铵态氮和硝态氮影响及其机理的研究还相对较少。本研究以黄土高原半干旱地区长期放牧的草地和围封7年的草地为对象, 分析土壤铵态氮和硝态氮的季节变化及其影响因素, 旨在为该区草地土壤质量评价、环境保护及制定合理的放牧强度等提供依据。
试验样地设在甘肃省榆中县的兰州大学半干旱气候与环境观测站(SACOL) (35° 57' N, 104° 09' E), 海拔1 965.8 m。该区气候类型属于大陆性半干旱气候, 为典型的黄土高原地貌, 年降水量约382 mm; 年平均气温6.7 ° C, 无霜期115 d, 年蒸发量917~1 368 mm; 年日照时数2 600 h[18, 19]。属于半干旱典型草原, 草地植物主要有本氏针茅(Stipa bungeana)、二裂委陵菜(Potentilla bifurca)、赖草(Leymus secalinus)、铁杆蒿(Artemisia gmelinii)、阿尔泰狗娃花(Heteropappus altaicus)、冷蒿(A. frigida)和茵陈蒿(A. capillaris)等。该地区草地原为传统耕作农田, 于1986年部分弃耕并恢复为天然草地[20]。
在观测站内选择两块地势平缓、坡向相近、面积各2 hm2的草地作为研究样地, 一块为围封草地, 该围封草地原为放牧草地, 围封开始时间为2005年10月[18]; 另一块为放牧草地, 放牧强度分别为冷季(11月至次年2月)1~2羊单位· hm-2, 暖季(3月至10月)2~3.5羊单位· hm-2。在围封、放牧草地中各随机设置3个50 m× 50 m的重复样区。
土壤样品于2013年5、6、7、8、9月每月中下旬进行采样, 8月份采集土样时同时用环刀法测定土壤容重。在围封与放牧草地每个重复样区中随机设置两条样线(间距为17 m), 沿每条样线等距离设置5个采样点, 采用土钻法分层采集0-10、10-20 cm土壤样品, 同一样线5个样点的样品混为1个混合样品。将土样中的植物根系及杂物去除后, 带回实验室进一步处理与分析。同时在每个重复样区内随机设置6个1 m× 1 m样方, 每个样地共计18个样方, 齐地刈割植被地上部分, 带回实验室后区分为活体(绿色生物量)和枯落物, 于105 ℃下杀青15 min后, 65 ℃烘干称重[21]。
8月采集的土壤样品用于土壤理化性质的测定。新鲜土壤样品过2 mm筛后, 用四分法分出一部分土壤样品, 带回实验室立即测定N-N、N-N和土壤微生物生物量碳。N-N采用氯化钾浸提-靛酚蓝比色法测定; N-N采用酚二磺酸比色法测定; 5、7、9月的土壤样品分取一部分, 采用氯仿熏蒸提取法[18]测定土壤微生物生物量碳; 所有土壤样品风干后过筛测定土壤有机碳、全氮、全磷及土壤pH值。土壤有机碳(SOC)采用重铬酸钾氧化滴定法测定; 土壤全氮和全磷的测定, 取过0.5 mm筛土样, 加10 mL硫酸, 加3.3 g催化剂(K2SO4∶ CuSO4· 5H2O=10∶ 1)消化, 然后置于流动注射分析仪测定(FIAstar 5000); 所有元素含量分析结果以烘干重表示(105 ° C); 土壤pH值采用2.5∶ 1水土比, 悬液用pHS23C型酸度计测定。土壤理化性质结果见表1。
| 表1 围封与放牧草地0-10和10-20 cm土层土壤理化性质 Table 1 Soil properties in 0-10 and 10-20 cm soil layers of fenced and grazed grasslands |
应用Excel软件进行数据处理, 采用Sigmaplot 12.5绘图, 应用SPSS 17.0进行方差分析, 应用Pearson相关性分析法分析了土壤铵态氮、硝态氮与土壤含水量、土壤微生物生物量碳及植被地上绿色生物量的相关性, 采用Canoco 4.5软件对土壤铵态氮、硝态氮与土壤含水量、土壤微生物生物量碳及植被地上绿色生物量关系进行冗余分析(RDA)并作图。
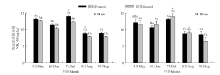
围封与放牧草地0-10和10-20 cm土层土壤铵态氮和硝态氮含量均表现出明显的季节变化(图1、图2)。
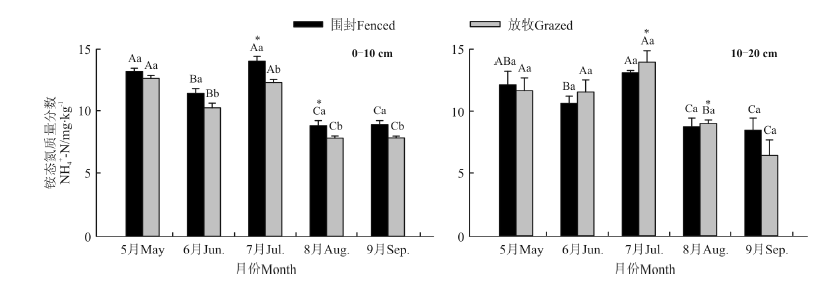 | 图1 围封与放牧草地不同土层土壤铵态氮含量的季节变化Fig.1 The seasonal variation of N-N content in different soil layers in fenced and grazed grasslands 注:不同小写字母表示同一土层同一月份两样地之间差异显著 (P< 0.05), 不同大写字母表示同一土层同一样地不同月份间差异显著(P< 0.05), “ * ” 表示同一处理不同土层之间差异显著 (P< 0.05), 下同。 Note: Different lower case letters mean significant differences between the two sites at 0.05 level, and different capital letters for the same plot mean significant difference within the same soil layer among different months at 0.05 level, “ * ” means significant difference between two soil layers in the same treatment at 0.05 level. The same below. |
 | 图2 围封与放牧草地不同土层土壤硝态氮含量的季节变化Fig.2 The seasonal variation of N-N contents in different soil layers in fenced and grazed grasslands |
季节尺度上, 围封与放牧草地0-10 cm土层土壤铵态氮含量分别为8.89~13.24和7.90~12.68 mg· kg-1, 硝态氮含量分别为1.12~3.36和0.77~2.97 mg· kg-1; 围封与放牧草地10-20 cm土层土壤铵态氮含量分别为8.48~13.10和6.48~13.95 mg· kg-1, 硝态氮含量分别为0.10~1.79和0.83~1.56 mg· kg-1。整个生长季中除了5月份, 围封草地0-10 cm土层铵态氮含量均显著高于放牧草地 (P< 0.05)。两样地10-20 cm土层铵态氮含量的季节变化呈现与0-10 cm土层相似的变化趋势, 除了7和8月份, 其它几个月两样地0-10和10-20 cm土层间铵态氮含量差异均不显著(P> 0.05); 整个生长季围封草地0-10 cm土层硝态氮含量均显著高于放牧草地; 两样地10-20 cm硝态氮含量的季节变化则均呈现单峰曲线的变化趋势, 且在不同季节均显著低于0-10 cm土层 (P< 0.05)。整个生长季, 10-20 cm土层铵态氮和硝态氮含量在围封和放牧草地之间均差异不显著(P> 0.05)。将围封与放牧草地不同时期各土层铵态氮与硝态氮含量进行相关分析得出, 除了围封草地10-20 cm土层外, 两草地不同土层铵态氮与硝态氮含量的季节变化之间均呈显著相关 (P< 0.01)(表2)。
| 表2 围封与放牧草地0-10和10-20 cm土层土壤铵态氮与硝态氮的相关关系 Table 2 Relationship between soil N-N and N-N in 0-10 and 10-20 cm soil layers in fenced and grazed grasslands |
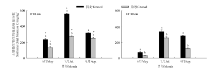
2.2.1 围封对土壤水分的影响 围封与放牧草地0-10和10-20 cm土层土壤含水量均在5月份以后逐渐升高, 于7月份达到最高值, 围封草地0-10 cm土层土壤含水量在5、6、9月份显著高于放牧草地 (P<
0.05), 而7、8月份两样地之间差异不显著(P> 0.05); 围封草地6月份10-20 cm土层的土壤含水量显著高于放牧草地, 而7月份放牧草地显著高于围封草地, 其它月份两样地间差异不显著。围封草地两土层土壤含水量在6、7月份、放牧草地两土层土壤含水量在6月份无显著差异, 其它月份两样地10-20 cm土层土壤含水量均显著高于0-10 cm土层(P< 0.05)(图3)。
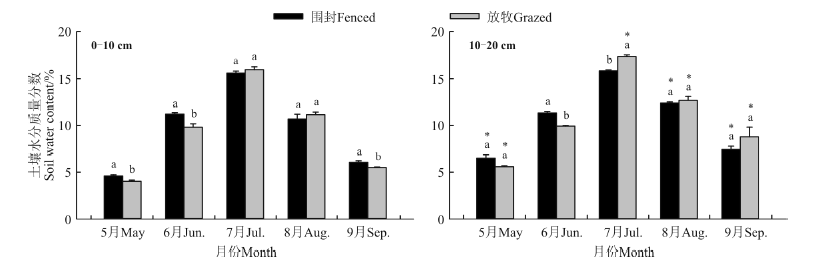 | 图3 围封和放牧草地0-10和10-20 cm土层土壤含水量季节变化Fig.3 Seasonal variation of soil water content in 0-10 and 10-20 cm soil layers in fenced and grazed grasslands |
2.2.2 围封对土壤微生物量碳的影响 2013年5月、7月和9月, 围封草地0-10和10-20 cm土层微生物生物量碳均显著高于放牧草地 (P< 0.05)(图4), 除9月份围封草地两土层之间差异不显著外(P> 0.05), 整个生长季两样地0-10 cm土层土壤微生物生物量碳均显著高于10-20 cm土层的(P< 0.05)。
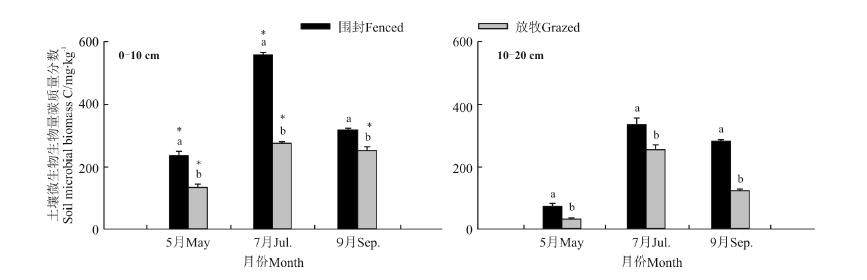 | 图4 2013年围封草地和放牧草地0-10和10-20 cm土层的土壤微生物量碳的季节变化Fig.4 Seasonal variations of soil microbial biomass C in 0-10 and 10-20 cm soil layers in the fenced and grazed grassland in 2013 |
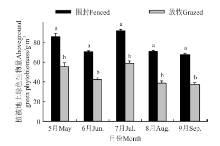
2.2.3 围封对植被地上绿色生物量的影响 植被地上绿色生物量与两土层铵态氮和硝态氮呈现相似的季节变化趋势(图5)。围封与放牧草地均在7月份出现峰值, 分别为91.50和58.70 g· m-2。在整个季节, 围封草地提高了植被地上绿色生物量(P< 0.05)。
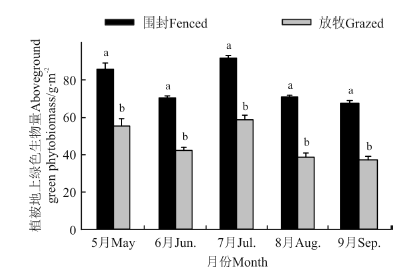 | 图5 围封和放牧草地植被地上绿色生物量的季节变化Fig.5 Seasonal variations of above-ground green biomass in the fenced and grazed grassland |
分别对0-10和10-20 cm土层中的铵态氮、硝态氮含量和土壤含水量、土壤微生物生物量碳及植被地上绿色生物量进行RDA分析, 以分析土壤含水量、土壤微生物生物量碳及植被地上绿色生物量对土壤铵态氮、硝态氮影响的程度。结果表明, 第一典范轴和所有典范轴的蒙特卡罗检验均呈极显著差异(P=0.002; P=0.002), 所以RDA排序结果是可靠的。这说明, 土壤含水量、土壤微生物生物量碳及植被地上绿色生物量可以很好地解释土壤铵态氮、硝态氮变化(表3)。
| 表3 土壤铵态氮、硝态氮和土壤含水量、土壤微生物生物量碳及植被地上绿色生物量的RDA分析结果 Table 3 Redundancy analysis results of soil N-N and N-N contents and soil water content, soil microbial biomass C, above-ground green biomass |
RDA分析结果也表明, 0-10 cm的4个排序轴特征值分别为0.718、0.006、0.267、0.008(表3), 其中第1轴是约束性排序轴, 对响应变量的解释比例达71.8%, 后3个轴是非约束性的, 4个排序轴的所有典范特征值之和占总特征值的72.4%。0-10 cm土壤铵态氮、硝态氮与土壤含水量、土壤微生物生物量碳及植被绿色生物量的相关性在前两个排序轴分别为0.854和0.658, 而10-20 cm的4个排序轴特征值分别为0.595、0.004、0.283、0.117, 4个排序轴的所有典范特征值之和占总特征值的60%, 且4个排序轴解释变量为99.9%, 10-20 cm土壤铵态氮、硝态氮含量与土壤含水量、土壤微生物量碳及植被绿色生物量之间的相关性在前两轴分别为0.826和0.187。整体来看, 0-10和10-20 cm的土壤铵态氮、硝态氮与土壤含水量、土壤微生物生物量碳及植被绿色生物量的RDA排序效果均较好。
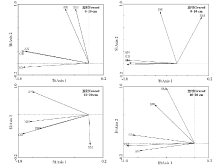
RDA排序结果仅能反映土壤铵态氮、硝态氮均与土壤含水量、土壤微生物生物量碳及植被地上绿色生物量间存在相关关系(箭头越长表示某一因子对土壤铵态氮、硝态氮的影响越大, 夹角越小相关性越高)(图6), 因此, 应用RDA分析中的前向选择和蒙特卡罗检验分析土壤含水量、土壤微生物生物量碳及植被地上绿色生物量对土壤铵态氮、硝态氮影响的显著性。结果表明, 0-10 cm土层围封和放牧草地RDA排序图中(图6), 植被地上绿色生物量对土壤铵态氮和土壤硝态氮响应有显著性解释(P=0.002; P=0.002); 而10-20 cm土层围封草地RDA排序图中(图6), 土壤含水量和植被地上绿色生物量均对土壤硝态氮有显著性解释作用(P=0.002), 而10-20 cm土层放牧草地植被地上绿色生物量对土壤铵态氮和硝态氮响应有显著性解释(P=0.002)。结果表明, 两土层土壤铵态氮和硝态氮均受植被地上绿色生物量影响, 而围封草地10-20 cm土层的土壤硝态氮还受到土壤含水量的影响。
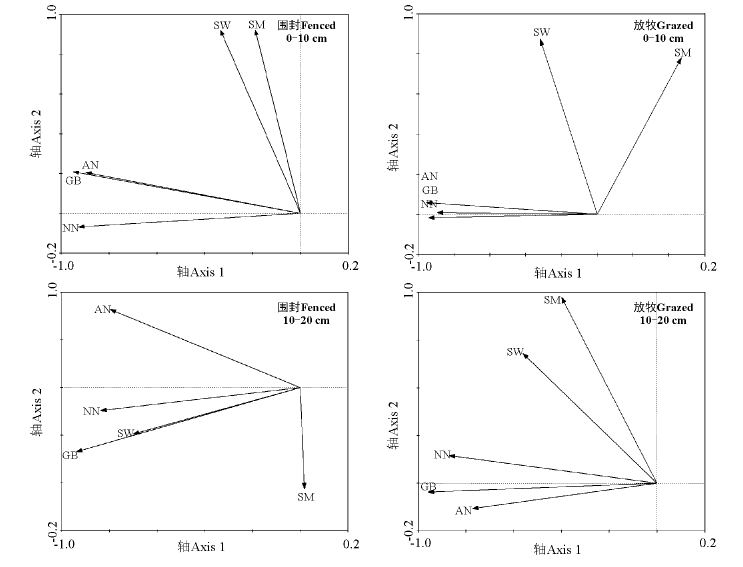 | 图6 围封与放牧草地不同土层土壤铵态氮、硝态氮与土壤含水量、土壤微生物生物量碳及植被地上绿色生物量的RDA排序图Fig.6 RDA ordination of N-N, N-N contents in different soil layers and soil water content, soil microbial biomass carbon, above-ground green biomass in fenced and grazed grasslands 注: AN, 土壤铵态氮; NN, 土壤硝态氮; GB, 植被地上绿色生物量; SW, 土壤含水量; SM, 土壤微生物生物量碳。 Note: AN, soil ammonium nitrogen; NN, soil nitrate nitrogen; GB, above-ground green biomass; SW, soil water content; SM, soil microbial biomass carbon. |
土壤中的有机氮通过微生物作用产生能被植物所吸收利用的有效氮(主要为铵态氮与硝态氮), 即氮素的矿化过程[22]。土壤中铵态氮和硝态氮主要来源于土壤中微生物的矿化作用, 包括硝化作用和氨化作用, 因此其含量的高低主要受土壤有机质含量、微生物种类、土壤环境条件以及人类活动(主要包括放牧、割草、施肥和火烧等)等因素的影响[23, 24]。基于本研究中所分析的指标及结果, 研究区草地利用方式的变化(放牧与围封)主要通过影响以下因素而对土壤铵态氮和硝态氮的含量产生影响。
土壤氮素含量及其有效性与土壤有机碳含量具有密切联系[23]。在黄土高原地区, 由于人类活动的干扰, 使得多数天然草地生产力大大降低, 加剧了水土流失与养分损失[25]; 有研究表明, 过度放牧导致天然草地生态系统退化, 生物多样性减少, 使适口性好、含氮量高的物种减少, 含氮量低的物种增加, 进而抑制氮素循环[24]。而草地围封后生物量显著增加, 一方面地表聚集了大量的枯落物, 使得以枯落物和根系分泌物的形式输入土壤的有机质增多, 提高了土壤全氮含量; 另一方面还可以提高土壤微生物活性, 增强土壤中氮素的固定和转化, 进而提高了土壤氮素的有效性[23, 26]; 此外, 植物残体在腐解过程中由微生物分解而产生的酸类物质, 可以使土壤中那些难溶性物质向有效性方向转化, 从而促进有效氮的增加[26]; 以上研究结果与本研究结果一致, 说明植被地上绿色生物量是影响上下两土层土壤铵态氮和硝态氮的主要因素之一。另有研究表明, 地表径流会导致氮流失, 而植被凋落物以及植株的密度、高度等均会对地表径流产生影响[17]。本研究中, 放牧草地围封后, 植被的高度和盖度增加, 有效地降低了风蚀和水蚀的程度, 进而减少了氮素的流失。
本研究中, 铵态氮与硝态氮呈极显著正相关(P< 0.01), 在其它研究中也发现这一现象。研究鼎湖山马尾松(Pinus massoniana)人工林时发现硝态氮和铵态氮在试验的大部分季节表现出显著的线性相关关系(P< 0.05)[3]。在哥斯达黎加的研究发现, 土壤铵态氮的供应会对土壤硝化速率产生影响[27], 亚马逊热带林和美国威斯康星州也发现土壤铵态氮和硝态氮之间具有正相关关系[28, 29]。本研究中, 整个生长季两样地土壤N-N 含量均显著高于N-N。有研究表明, 植物从土壤中吸取养分供自身生长时会优先利用土壤中的硝态氮, 从而使土壤的硝态氮净剩余量减少[23, 24, 25, 30], 另外, 相较于带正电荷的N-N, 带负电荷的N-N更容易通过淋溶作用而损失, 使得生态系统中N-N浓度较高, 并以此来保持土壤中氮素的含量[31], 这可以一定程度上解释本研究区土壤中铵态氮含量高于硝态氮的原因。本研究中, 围封和放牧草地0-10 cm土层的铵态氮与硝态氮含量在5月份较高, 可能是因为非生长季铵态氮与硝态氮的累积效应, 而刚刚返青的植物生长速率较慢, 对养分的需求量较低, 从而使其含量维持在较高水平[30]; 6月份植物进入快速生长阶段, 增加了对氮素的吸收利用, 使两样地0-10 cm土层的铵态氮与硝态氮含量均明显降低。
土壤水分是影响土壤微生物生长、发育和繁殖的重要环境因子之一[32], 并通过控制微生物的生长和活性而进一步影响土壤的氮矿化过程[33]。围封与放牧对土壤水分的影响比较复杂, 一方面, 与围封草地相比, 放牧草地由于家畜的采食和践踏, 增加了土表所接受到的太阳辐射和土表裸露面积, 降低了植被盖度和植被地上绿色生物量, 增加了土壤温度和土壤水分的蒸发损失, 导致土壤含水量降低。另一方面, 围封后, 植被地上绿色生物量和植被盖度增加, 使得植物蒸腾速率加快, 提高了对土壤水分的利用, 导致土壤含水量降低。因此, 围封与放牧对土壤含水量的影响取决于植物对土壤水分的利用效率和土壤水分的自身蒸发速率这两个过程之间的平衡[18]。在一定范围内, 土壤净氮矿化速率和净硝化速率均随着土壤水分的升高而增加[15, 34]。本研究中, 0-10 cm土层土壤铵态氮、硝态氮含量在大多数季节中表现为围封草地显著高于放牧草地, RDA冗余分析结果表明, 土壤水分对围封和放牧草地10-20 cm土层土壤硝态氮含量有显著的影响, 说明对于半干旱区草地, 土壤水分也是影响土壤氮素矿化的重要因素之一。另有研究表明, 硝态氮会受到土壤含水量影响的原因可能是土壤水分可以调节硝化细菌种群数量[35], 而且, 在一定的土壤含水量范围内, 土壤水分的增加有利于硝化细菌的生长, 而不利于反硝化细菌, 使得铵态氮的固持小于硝化, 进而使硝态氮含量增加[34]。本研究中, 围封草地0-10 cm的土壤含水量在多数季节显著高于放牧草地(P< 0.05), 这可能是围封草地增加了土壤硝态氮的原因之一。
土壤氮素的矿化和硝化作用受微生物的影响, 微生物参与并调控其过程, 同时, 土壤微生物量是土壤氮矿化的重要氮源, 也是组成土壤有效态氮的重要部分[31]。草地土壤的微生物数量通常与土壤养分和土壤水分之间存在不同程度的正相关关系[32], 而植物根系的代谢活动和植物枯落物对微生物(即分解者)的生存环境也会产生影响[36, 37], 并对土壤有机质的分解产生显著的影响。放牧活动会引起土壤微生物量碳的变化, 可能是由于长期的放牧促进了某些微生物种群的生长, 而抑制了其它微生物种群的生长, 导致土壤微生物功能多样性指数下降[35]; 另外, 放牧干扰下, 牲畜的践踏改变了土壤结构, 降低了土壤透水性、透气性, 使得土壤微生物的繁殖代谢受到强烈的干扰, 进而降低了氮的矿化速率[38], 使得放牧草地土壤铵态氮和硝态氮显著低于围封草地。围封有助于土壤微生物的恢复[30, 39], 这是由于围封一方面为土壤氮素矿化提供了物质和能量, 另一方面为微生物提供了较多的能源与养分, 促进微生物大量繁殖及新陈代谢, 有利于提高微生物群落物种数量和多度, 从而促进了土壤氮素的矿化[35]。本研究中, 草地围封后植被得以恢复, 使土壤有机碳含量显著增加(表1), 加之围封后多数季节土壤水分的增加, 使得围封草地土壤微生物生物量碳显著高于放牧草地。然而冗余分析结果显示, 土壤微生物生物量碳并没有对两样地土壤铵态氮和硝态氮产生显著影响。一方面, 可能是因为5月份围封和放牧草地0-10 cm土层较高的铵态氮与硝态氮含量并非当季土壤氮素矿化的结果, 另一方面, 微生物生物量碳测定的频次相对较低, 从而可能会对冗余分析结果造成影响。因此, 关于该研究区微生物生物量碳对土壤氮素矿化的影响还有待更深入的研究。
围封与放牧草地0-10和10-20 cm土层土壤铵态氮和硝态氮含量均表现出明显的季节变化, 且多数季节围封草地0-10 cm土层铵态氮和硝态氮含量均显著高于放牧草地 (P< 0.05); 两样地土壤铵态氮与硝态氮之间存在极显著正相关(P< 0.01), 且铵态氮含量显著高于硝态氮含量。围封与放牧草地0-10和10-20 cm土层土壤铵态氮和硝态氮均受植被地上绿色生物量的影响, 而围封草地10-20 cm土层土壤硝态氮还受土壤水分的影响; 放牧草地围封后, 植被生物量及土壤微生物生物量碳显著增加, 并一定程度上增加了土壤水分, 这些均有利于土壤氮素的矿化。
The authors have declared that no competing interests exist.
| [1] |
|
| [2] |
|
| [3] |
|
| [4] |
|
| [5] |
|
| [6] |
|
| [7] |
|
| [8] |
|
| [9] |
|
| [10] |
|
| [11] |
|
| [12] |
|
| [13] |
|
| [14] |
|
| [15] |
|
| [16] |
|
| [17] |
|
| [18] |
|
| [19] |
|
| [20] |
|
| [21] |
|
| [22] |
|
| [23] |
|
| [24] |
|
| [25] |
|
| [26] |
|
| [27] |
|
| [28] |
|
| [29] |
|
| [30] |
|
| [31] |
|
| [32] |
|
| [33] |
|
| [34] |
|
| [35] |
|
| [36] |
|
| [37] |
|
| [38] |
|
| [39] |
|