

{kind=link}
{kind=link}
{kind=link}
{kind=link}
刈割频度和刈割时间对冷地早熟禾生长的影响
[马银山1  , 徐琳
, 徐琳1 , 范海鹏1 , 常立江1 , 周睿1 , 张世挺2 ]
, 徐琳|
|


在野外条件下,采用析因设计,对刈割频度和刈割时间变化下冷地早熟禾( Poa crymophila)生长情况进行研究,探讨冷地早熟禾对环境干扰的适应对策。刈割频度分为刈割1次、刈割2次和刈割3次,刈割时间分为早期刈割和晚期刈割。结果表明,所有刈割处理下,冷地早熟禾均发生了超补偿反应,超补偿的发生与刈割频度和刈割时间无关。地上生物量与分蘖数呈显著的正相关,冷地早熟禾是通过长出更多的分蘖来抵抗环境干扰的,新生分蘖产生所需的资源主要是由根部供应。冷地早熟禾对于不同时间的刈割处理,在生活史对策方面进行了权衡。早期刈割处理中,植物采取尽快产生种子,保证能够用种子繁殖后代的策略;晚期刈割处理中,植物采取了减少种子产生,保证植株个体来年存活和返青的策略。资源在植物体内进行重新分配和权衡是冷地早熟禾应对外界环境干扰的主要的适应对策。
With a purpose for strategies of Poa crymophila to adapt to environmental interference, we studiedthe growth of P. crymophila subject to varying moving frequencies and time using factorial design and in field conditions. Mowing frequencies are divided into one time, two times, and three times; and the mowing time is divided into early mowing and late mowing. The results showed that overcompensation effects occurred with all the P. crymophila in all cases, and the occurrence of overcompensation was irrelevant to moving frequencies and time. Above-ground biomass and tiller number had a significant positive correlation, P. crymophila resisted environmental interference by growing more tillers, and the resources required to produce tillers were mainly supplied by the roots. The mowing treatments of P. crymophila at different time were evaluated in respect of life history strategy. In case of early moving treatment, plants produced seeds as soon as possible to ensure their capability of production of the seeds; in case of late mowing, plants reduced the production of seeds to ensure the survival of individual plants in the coming year and taking root. Redistribution and balancing of resources in plants was the major strategy of P. crymophila to respond to interference of external environment.
许多研究表明, 中度放牧或刈割影响植物的生长, 其中刈割频度和刈割时间是影响植物生长很重要的因素。Dyer[1]和McNaughton[2]的放牧优化假设, 揭示了植物对食草作用的反应。假设指出, 中等强度的放牧或刈割刺激植物的生长。一些研究证明, 刈割或放牧对植物的生长有促进作用; 但另一些研究证明这种促进作用是有限的, 而且是有争议的[3, 4]。通常来说, 植物对植株部分移除有两种可能的反应:一方面, 它们尽可能通过补偿来替代损失的部分, 另一方面将资源重新进行分配或者进行转换; 对这两个方面如何选择, 主要受外界环境因素、植物预期寿命和刈割时间的影响[5, 6]。植物去除部分叶片后生物量增加, 称之为超补偿生长。落叶时间、频度、植物体损伤程度以及相邻植物竞争作用对植物补偿能力影响很大[7]。Hilbert等[8]研究认为, 生长缓慢的植物具有更强的补偿能力; 与之相反的是, Maschinski和Whitham[9]认为, 在资源丰富的条件下, 快速生长的植物, 将表现出更大的补偿能力; 但也有一些研究证实, 植物生长快慢与补偿能力大小并无直接关系[10, 11]。
植物对刈割或落叶的反应表现在叶面积或生物量的减少, 且是一个复杂的过程[12, 13]。植物在刈割或放牧中获益, 可能的机制主要有两种:1)与邻近高秆植物遮阴的影响去除有关。通过刈割或放牧, 去除了植物中死去的部分或减少了自身遮阴的光照竞争, 而使植物得到补偿。2)刈割能刺激光合作用, 促进植物生长。移除老的或死去的组织, 平衡了植物生物量的分配, 增加了生长率, 促进植物生殖分蘖增多, 出现了补偿[14, 15, 16, 17, 18]。
另外, 植物本身的特性决定了植物资源的分配格局[19, 20]。植物在食草作用下优先将资源分配给生长、贮藏、防卫或繁殖[21, 22]。根据这个优先原则, 随植物的生长, 补偿能力增加的原因可能主要是由于植物体将资源大量用于贮藏或者对抗食草作用的影响。生长和繁殖通常是竞争关系, 植物或许在生长和繁殖方面有一个权衡[23, 24]。而且, 繁殖对策主要有两种可能:植株在当年存活和生产种子繁殖后代这两方面的权衡有可能导致了补偿反应[19]。
放牧研究中, 多集中于放牧强度对植物生长影响的研究, 而关于放牧频度和刈割时间对植物生长影响的研究相对较少。放牧优化假说指出, 在放牧压力下, 中度放牧能促进植物生物量的增加, 发生超补偿。基于这个前提, 中等强度的多次放牧是否也会促进植物生物量的增加, 放牧频次的多少对超补偿有无影响?本研究通过对物种在中等强度下的刈割频度和刈割时间来验证这个假说。在放牧实践中, 牲畜放牧与青干草的收获往往存在矛盾, 牲畜放牧的时间和频次会影响青干草的收获, 反之青干草的收获早晚也会影响牲畜放牧的质量, 因此, 选择研究的牧草需具备既可进行牲畜放牧, 又可进行青干草储存的特点。本研究所选择的物种为亚高山草甸的优势牧草冷地早熟禾(Poa crymophila), 符合这个要求。本研究主要解决以下问题。1)刈割频度和刈割时间对牧草生长的影响是什么? 2)刈割或刈割频度是否影响超补偿的发生?3)冷地早熟禾对环境干扰的适应对策是什么?通过本研究, 探讨牧草对刈割频度和刈割时间的响应情况, 揭示牧草在刈割作用下发生超补偿反应的机制和原理, 旨为放牧草地合理控制放牧次数及选择合理的放牧时间提供理论参考。
试验地点位于甘肃省张掖市肃南康乐草原站, 地理位置为38° 8' N, 99° 6' E, 平均海拔3 200 m, 年均温3.6 ℃, 年均降水量 254.8 mm。
冷地早熟禾是疏丛型多年生草本, 茎秆直立, 柔软, 略带甜味, 适口性好, 根茎发达, 分蘖能力强, 是饲用价值较高的一种牧草。夏、秋是各类家畜抓膘的优良牧草、冬春则是救命保膘草, 青干草是冬春的良好补充饲草。种植当年实生苗只能达到孕穗期, 不能结实。在第2年4月下旬至5月上旬返青, 5月中旬至6月上旬孕穗, 6月上旬至7月上旬抽穗开花, 8月下旬种子成熟。在海拔2 300~4 400 m的高寒牧区, 均能生长良好, 是当地的优良牧草[6]。
试验采用析因设计, 两个因子为刈割时间和刈割频度。试验于2011年5月份开始进行种植, 种植后连续观察生长两年的数据。在野外, 平整一块25 m2的试验地, 划分为21个样方, 每个样方大小为1.3 m× 0.8 m, 样方中间留25 cm的过道。对21个样方的土壤进行深翻, 平整, 去除其它杂草。在每个样方均匀播撒80粒冷地早熟禾种子, 根据出苗情况进行间苗、移苗, 确保每个样方有50株个体, 且个体之间有一定距离(约12 cm), 避免植株个体之间的密度制约。苗期定期浇水, 保证幼苗成活[25]。
由于冷地早熟禾当年不能结实, 在种植的第1年只在收获时进行了生物量测定。第2年6月上旬开始, 根据当地夏季牧场和秋季牧场初次放牧的实际情况, 确定不同的初次刈割时间, 分为早期刈割(拔节期)和晚期刈割(抽穗期), 分别在两个不同时期开始, 按不同刈割频度(刈割1次、刈割2次和刈割3次)进行刈割处理, 刈割时对植物整株地上部分按高度进行50%的刈割处理, 模拟中度放牧。刈割频度的时间间隔为20 d, 每个处理重复3次, 有3个样方不进行刈割处理, 作为对照。在第一次刈割处理开始时, 每个样方中随机定株10株, 然后对整个样方的植株进行刈割处理, 以避免其它个体对定株个体遮阴的影响。定株个体所剪下的部分分为茎、叶、穗, 在85 ℃的烘干至恒重, 称重, 以计算累积地上生物量。整个试验在植物枯黄期进行收获, 包括对照样方, 从每个样方的定株个体中选取5株(每个处理共15株个体), 连根挖起, 清洗干净, 数出每株的分蘖数。将个体分为根、茎、叶、穗四部分在85 ℃的烘箱中烘到恒重, 用1/10 000的电子天平称重[25, 26, 27]。
收获后, 对每个个体分别计算累积地上生物量、根冠比(根生物量/累积地上生物量)及资源分配参数(繁殖生物量/累积地上生物量)。采用多因素方差分析确定刈割时间、刈割频度对累积地上生物量、根冠比、分蘖数和补偿指数的影响效应。为了使数据符合正态分布和改善方差齐性, 累积地上生物量、根冠比的数据均进行了常用对数(log10)转换。用QQ-Pots检验数据正态性, Levene’ s 检验法检验方差齐性(在显著性水平为0.05时不能拒绝数据的方差齐性假设); 用多重比较中的最小显著差数法( LSD method)或 Tamhane’ s 检验法(方差不齐时)比较各处理间差异。所有分析均用SPSS 13.0统计软件进行[28]。
刈割时间对地上生物量和分蘖数影响显著(P< 0.05), 但对根冠比影响不显著(P> 0.05)(表1); 刈割频度对地上生物量和根冠比影响极显著(P< 0.01), 对分蘖数影响显著(P< 0.05), 两因子之间的交互作用对地上生物量、分蘖数、根冠比均有显著影响。刈割时间只对茎分配有显著影响(表2), 对根分配、叶分配、繁殖分配并没有显著影响; 刈割频度只对根分配有极显著的影响, 对茎分配、叶分配、繁殖分配并没有影响; 两因子的交互作用对根分配和繁殖分配分别有显著和极显著影响。
| 表1 刈割频度和时间对冷地早熟禾生长参数影响的方差分析结果 Table 1 Results of applying ANOVA to study the effect of clipping frequency(CF) and clipping time(CT) on growth variables |
刈割时间和刈割频度显著影响冷地早熟禾的分蘖数(表1)。在刈割1次时, 晚期刈割的分蘖数显著大于早期刈割的(P< 0.05); 刈割2次、3次的处理下, 分蘖数在刈割时间之间差异不显著(P> 0.05)(图1)。早期刈割, 随着刈割频度的增加, 分蘖数无显著变化; 晚期刈割, 随着刈割频度的增加, 分蘖数出现先增加后减少的情况, 刈割1次和刈割2次的处理下分蘖数显著大于对照和刈割3次的处理。
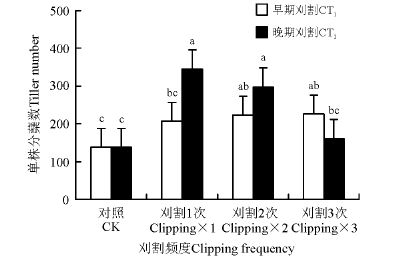 | 图1 刈割频度和时间对冷地早熟禾每株分蘖数的影响Fig.1 Effects of change of clipping frequency (CF) and clipping time (CT) on tiller number per plant 注:不同小写字母表示不同处理间差异显著(P< 0.05)。CT1表示早期刈割(6月), CT2表示晚期刈割(7月)。下同。 Note: Different lower case letters indicate significant difference between different treatments at 0.05 level. CT1, first clipping time in June, CT2, first clipping time in July. The same below. |
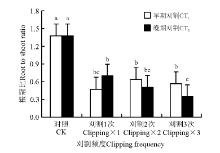
刈割使冷地早熟禾根冠比显著减小, 刈割和对照相比, 根冠比均显著小于对照(P< 0.05)(图2)。早期刈割时, 在3个刈割频度下, 根冠比均无显著变化(P> 0.05)。晚期刈割时, 刈割1次和刈割2次根冠比无显著差异; 但刈割1次显著高于刈割3次的根冠比(P< 0.05), 刈割3次的根冠比也显著小于对照的。
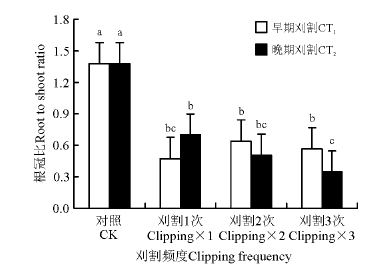 | 图2 刈割频度和时间对冷地早熟禾根冠比的影响Fig.2 Effects of change of clipping frequency(CF) and clipping time(CT) on root shoot ratio |
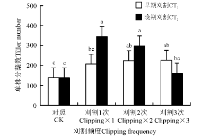
刈割可显著促进冷地早熟禾累积地上生物量的增加, 刈割累积地上生物量均显著大于对照(P< 0.05), 无论早期还是晚期刈割, 在各刈割频度下, 冷地早熟禾均发生了超补偿(图3)。说明对于冷地早熟禾来说, 中度刈割处理时, 均会发生超补偿, 而与刈割频度无关。早期刈割时, 累积地上生物量在刈割频度之间无显著差异(P> 0.05); 晚期刈割时, 刈割1次的地上生物量显著大于刈割3次的。刈割1次时, 晚期刈割显著大于早期刈割的累积地上生物量。刈割2次和3次时, 刈割早期和晚期差异不显著。
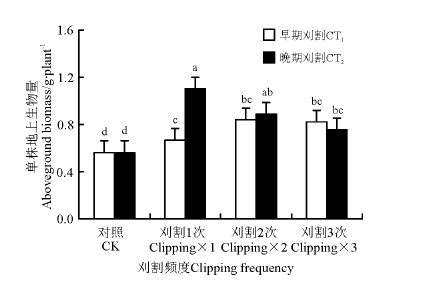 | 图3 刈割频度和时间对冷地早熟禾累积地上生物量的影响Fig.3 Effects of change of clipping frequency (CF) and clipping time (CT) on accumulated aboveground biomass |
刈割处理后植物显著减少了根的分配(P< 0.05)(图4A), 显著增加了茎的分配(图4B)。刈割1次和刈割3次处理相比, 晚期刈割茎分配显著大于早期刈割的。刈割处理下叶分配明显增加, 早期刈割, 刈割1次叶分配显著大于刈割2次的; 晚期刈割, 刈割2次和刈割3次显著大于对照, 但刈割1次与对照差异不显著(P> 0.05)(图4C)。刈割时间和刈割频度对冷地早熟禾繁殖分配无显著影响, 但两因子的交互作用影响显著(表2)。早期刈割1次时, 繁殖分配较对照无显著变化, 晚期刈割1次时, 繁殖分配较对照显著降低, 刈割2次和3次之间, 无论早期与晚期刈割, 繁殖分配之间无显著差异, 但都显著小于对照(图4D)。
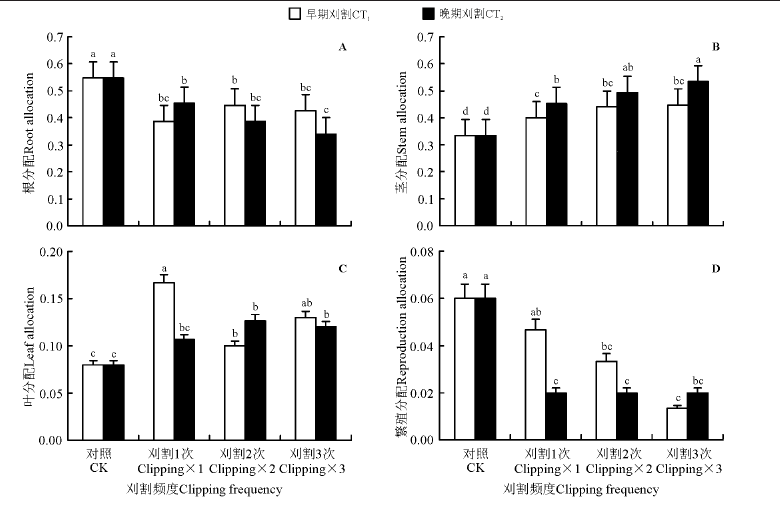 | 图4 刈割频度和时间对冷地早熟禾资源分配的影响Fig.4 Effects of change of clipping frequency (CF) and clipping time (CT) on resource allocation |
| 表2 刈割频度和时间对冷地早熟禾资源分配影响的方差分析结果 Table 2 Results of applying ANOVA to study the effect of clipping frequency(CF) and clipping time(CT) on resource allocation |
食草作用或放牧通过以下4种可能机制改变植物资源利用效率和植物生长特性:1)植物幼年阶段光合作用的维持导致营养物质在叶上的累积; 2)刺激植物生长导致生物量的增加; 3)通过食草作用刺激分蘖, 增加茎的密度; 4)通过食草作用的选择, 减少死去的生物量, 导致植物体组成部分向不同方面的转换[29, 30, 31]。
van der Graaf等[32]认为, 第2和第3个机制更切合实际; 但对这两个机制, 也有不一致的研究结果[33, 34]。许多研究表明, 移除顶芽能刺激牧草的分蘖, 中度落叶有可能打破了顶端优势, 刺激被刈割后的植物长出更多的分蘖[9]。但有些研究也得出与之相反的结论[35, 36]。 在本研究中, 冷地早熟禾在所有刈割处理中都出现了分蘖增加的情况, 这个结果与移除顶芽能刺激牧草分蘖和研究结果相一致。
在植物体中, 子蘖的营养供给在一定程度上依靠母蘖, 当植物受到环境条件压力时, 子蘖由于竞争资源困难而首先死亡, 但资源充足时, 分蘖的生长不受影响[37]。冷地早熟禾在刈割处理后, 由于大量遮阴的叶片被清除, 光照强度增大, 光照资源的增加可以为植物提供部分所需的资源供应。从表3可知, 根生物量与分蘖数呈极显著的正相关(r=0.664, P< 0.01), 根分配与分蘖数呈显著的正相关(r=0.200, P< 0.05), 这说明, 分蘖数的增加与根部资源储存和分配的多少有关。一方面根部储存的资源可以直接供给新生分蘖的生长, 另一方面光合作用产生的资源, 有少量分配给根, 再由根供给了新生分蘖的生长。从根分配和根冠比也可以看出, 根部资源出现了从地下部分向地上部分转移的情况, 根部资源向地上部分转移过程中, 首先为新生分蘖提供了大量的营养, 新生分蘖资源供应充足, 只有少量的死亡, 与死亡的分蘖数相比, 增加数量大于减少数量, 所以分蘖数出现了增加。分蘖数与繁殖分配呈极显著负相关(r=-0.613, P< 0.01), 虽然分蘖数增加了, 但繁殖分配却总体出现了减少的情况, 这可能是由于新生分蘖的资源供应和繁殖部分的资源供应来源不同而导致了这样的结果。新生分蘖的资源可能主要由根部供应, 而繁殖枝生长的资源主要由光合作用供应。冷地早熟禾在生长过程中, 在受到刈割等外界干扰时, 有可能为了保证个体或子代的延续, 放弃了开花结实这一比较耗费资源的生活史对策, 而将少量光合作用资源用于新生分蘖。
| 表3 各生物量参数之间的相关分析 Table 3 Results of correlation analysis between each growth parameters |
在植物体中, 生物量移除表现出补偿反应是常见的现象[38]。但刈割后的超补偿现象并不是在所有植物体中都出现。冷地早熟禾在中度刈割时, 所有的刈割频度处理中都出现了超补偿情况, 从表3可以看出, 冷地早熟禾分蘖数与地上生物量呈极显著正相关(r=0.788, P< 0.01), 这说明, 中度刈割处理下, 冷地早熟禾通过长出更多的新生分蘖来弥补叶的损失, 分蘖数的增多导致地上生物量的增加。另外, 地上生物量与根冠比呈显著负相关(r=-0.058, P< 0.05), 在所有的刈割处理中, 与对照相比, 根冠比都减小, 地下部分资源分配减少, 茎和叶的资源分配显著增加。资源从根到茎的再分配可以很好地解释中度刈割后植物的反应[39]。植物体部分组织被移除的补偿, 尤其是大量落叶后, 需要很多的资源补充[33, 40]。植物体茎、叶等部分被移除, 导致了植物失去了很多的光合组织, 刈割有可能改变了植物资源分配模式, 很多研究也证实了这一点[41, 42]。本研究中, 冷地早熟禾在地上部分刈割后, 通过利用地下部分储藏的能量转移到地上部分, 促进分蘖的生长。虽然植物体有大量的落叶, 光合作用能力减弱, 资源减少, 但地下部分资源向地上部分的重新分配过程中, 为新生分蘖首先提供了营养。新生分蘖长出后, 通过植物光合作用能力的提高来恢复植物体资源的供应, 再反过来促进了植物的生长, 导致了植物地上生物量的增加, 出现了超补偿。
刈割时间在超补偿中是一个重要的因子[43, 44, 45]。但是在本研究中, 两个刈割时间之间, 超补偿反应并没有显著差异。本研究初次刈割时间是根据青藏高原亚高山草甸的当地夏季牧场和秋季牧场开始放牧时间选择的。由于早期和晚期刈割时间间隔一个月, 这两个时间段植物生长处于不同的生长阶段, 刈割后植物不同的生活史对策可以很好地解释两个刈割时间之间超补偿反应没有显著差异的问题。资源或许在植物体内部朝着不同的生活史功能进行分配[46], 依赖于环境的变化、竞争的优势或植物生长的模式[47]。早期刈割时, 植物处于拔节期, 刈割去除的主要是茎和叶, 植物的光合作用受到影响, 光合作用降低, 但在这个阶段, 植物地上生物量的增加一部分是靠地下部分的资源转移, 另一部分是靠光合作用产生的资源, 光合作用产生的资源主要用于繁殖部分的生长, 植物在此时选择的生活史对策是尽快生产种子, 保证植物能够利用种子进行子代的繁殖。晚期刈割时, 植物处于抽穗期, 繁殖枝的生长已经基本结束, 刈割去除了部分的繁殖枝, 导致植物种子生产的过程受到影响, 植物已经没有更多资源用于繁殖枝的再生, 而且产生繁殖枝的成本太高, 此时植物选择了对资源的重新分配。光合作用的资源除了用于部分种子生产外, 一部分向地下部分运输, 储藏在根中, 以利于来年植株的返青。晚期刈割时, 植物在产生种子和来年返青两个生活史对策方面进行了权衡, 降低了植物个体死亡的风险。所以, 在早期刈割和晚期刈割虽然都发生了超补偿, 但补偿的程度差异不显著, 植物在繁殖对策的选择中进行了权衡。资源充足时尽可能进行种子繁殖, 资源不足时, 选择种子繁殖和营养繁殖两种方式, 以保证子代的延续。
对于冷地早熟禾来说, 中度刈割情况下, 刈割频度和刈割时间对超补偿的发生没有影响, 刈割3次仍发生超补偿, 超补偿的发生主要是分蘖数增多引起的。冷地早熟禾对于不同时间的刈割处理, 在生活史对策方面进行了权衡。早期刈割处理中, 植物采取尽快生产种子从而有足够种子繁殖后代的策略; 晚期刈割处理中, 植物采取了减少种子生产, 保证植株个体来年存活和返青的策略。资源在植物体内进行重新分配和权衡是冷地早熟禾应对外界环境干扰的主要的适应对策。
The authors have declared that no competing interests exist.
| [1] |
|
| [2] |
|
| [3] |
|
| [4] |
|
| [5] |
|
| [6] |
|
| [7] |
|
| [8] |
|
| [9] |
|
| [10] |
|
| [11] |
|
| [12] |
|
| [13] |
|
| [14] |
|
| [15] |
|
| [16] |
|
| [17] |
|
| [18] |
|
| [19] |
|
| [20] |
|
| [21] |
|
| [22] |
|
| [23] |
|
| [24] |
|
| [25] |
|
| [26] |
|
| [27] |
|
| [28] |
|
| [29] |
|
| [30] |
|
| [31] |
|
| [32] |
|
| [33] |
|
| [34] |
|
| [35] |
|
| [36] |
|
| [37] |
|
| [38] |
|
| [39] |
|
| [40] |
|
| [41] |
|
| [42] |
|
| [43] |
|
| [44] |
|
| [45] |
|
| [46] |
|
| [47] |
|