{kind=link}
芜菁甘蓝种质资源评价
[王应芬1, 2 , 牟琼1  , 李娟
, 李娟1 , 雷霞1 , 吴佳海1 ]
, 李娟|
|


第一作者:王应芬(1977-),女,贵州湄潭人,高级畜牧师,硕士,主要从事草地建植及养殖研究。E-mail:[email protected]
为有效利用芜菁甘蓝( Brassica napobrassica)种质资源,对28份芜菁甘蓝种质资源的27个数量性状进行了观测,并分析其遗传多样性。结果表明,1)不同材料间存在较大差异,且不同材料在不同性状上也表现出不同程度的多样性。其中,单株角果数的变异程度最大,变异系数达64.1%;其次为小裂叶对数,其变异系数为57%;单根重的变异系数最小,为2.4%。2)通过聚类分析,将28份芜菁甘蓝分为四大类群。其中第Ⅰ类的14份材料均具有绿色子叶、侧芽小、板叶叶型、叶顶端圆形、根形椭圆及须根少等特点;第Ⅱ类中的两份材料具有平滑叶面、叶面无刺毛、少蜡粉、叶面有光泽及半直立叶簇等特点;第Ⅲ类由分蘖能力中等、浅绿色角果、白色肉质根及少须根的6份材料组成;第Ⅳ类由具有复锯齿和大波纹叶缘、有叶瘤及红褐色种皮的6份材料组成。
In order to efficiently utilize Brassica napobrassia, the genetic diversity of 28 B. napobrassica accessions were assessed by measuring 27 quantitative traits. The results showed that the B. napobrassica had significant genetic differentiation and diversity in 27 quantitative traits. Among those data, the coefficient of variation (CV) of the nutlet was maximum, reaching up to 64.1%, the second was the pair number of lobule with CV of 57%, and the minimum was weight per earthnut with CV of 2.4%. In addition, 28 B. napobrassica accessions were divided into four categories by cluster analysis. The first group included 14 accessions possessingthe characteristics of green cotyledon, small lateral bud and round blade leaf opex, the second group included 2 accessions with the characteristics of smooth leaf surface, no seta on leaf surface, less powder and semi upright leaf cluster, the third group included 6 accessions with the characteristics of medium tillering ability, light green siliqua, white fleshy root, and less fibrous root, the fourth group included 6 accessions with the characteristics of serrated and large corrugated leaf edge, leaf nodule and reddishbrown seed coat.
芜菁甘蓝(Brassica napobrassica), 又称洋大头菜、洋蔓菁及洋疙瘩等。芜菁甘蓝为芜菁(B. rapa)和甘蓝(B. oleracea)杂交形成, 最初来源于地中海沿岸及瑞典等地, 在我国主要栽培于贵州、云南、山东、河北等省(区), 现在江西、湖南、广西、江苏等省(区)也有推广种植。自芜菁甘蓝出现后, 牧场主们发现它比芜菁更富有营养价值, 被誉为是一种具有广泛推广发展前途的优质高产块根饲料作物[1]。
芜菁甘蓝块根肥大, 产量高, 且肉质根干物质含量高, 尤其是糖分、淀粉含量多, 且维生素C含量也较丰富[2]。芜菁甘蓝以肥大的肉质根为主要产品, 茎叶均可利用, 主要用于秋季采叶青饲, 冬、春季采收肉质根和茎叶饲喂畜禽, 可缓解冬、春季青绿饲料缺乏的矛盾。芜菁甘蓝汁多味甜, 适口性好, 营养丰富、产量高; 适于切碎后直接饲喂或青贮、打浆后利用, 猪、牛、羊等家畜喜食[3, 4]。
目前, 芜菁甘蓝的种植以常规品种为主, 存在纯度、块根一致性差, 根型、根皮色参差不齐等问题。同时, 块根用芜菁甘蓝种质资源的遗传背景较为狭窄、育种材料较单一, 严重制约了芜菁甘蓝的生产和推广及根用芜菁甘蓝的育种水平[5]。因此, 本研究通过观测28份芜菁甘蓝种质的27个植物学性状表现, 分析其遗传多样性, 旨为进一步开展芜菁甘蓝种质资源分子水平的遗传多样性和相关性研究提供理论指导, 同时为我国根用芜菁甘蓝资源的组成结构及优势育种工作提供基础数据。
试验材料为在贵州各地和其它地区采集到的地方品种资源, 共28份。主要包括当地原有品种和长期在当地栽培过的引进种。其中, 贵州9份, 省外12份, 国外7份。命名方式为有名字的品种按照当地称谓命名, 无名字的品种或存在同名现象的品种, 以采集地的乡镇名字命名(表1)。
| 表1 供试材料名称与来源 Table 1 Names and origins of Brassica napobrassica germplasm resources in this experiment |
试验设在贵州省草业研究所试验基地, 位于107° 33' E, 25° 52' N, 海拔960 m, ≥ 10 ℃年积温4 538 ℃· d, 极端低温-8 ℃, 极端高温38 ℃, 最高月平均气温27.6 ℃, 年均温15 ℃, 年降水量1 346.3 mm, 降水多集中在5-8月, 无霜期272 d, 年日照时数1 336.7 h, 空气相对湿度82%, 雨热同期, 干湿季节较明显。试验地土壤为黄壤, 有机质含量2.8 g· kg-1, 全氮含量0.85 g· kg-1, 有效磷含量30.62 mg· kg-1, 速效钾含量86.2 mg· kg-1, pH值6.5。前茬作物为多花黑麦草(Lolium multiflorum)。
1.3.1 播种 播种前犁耙两次, 并辅以人工碎土平整。基施腐熟农家肥2.25 kg· m-2和复合肥74.96 g· m-2。穴播, 每穴播种5~8粒种。9月5日播种, 顺序排列, 小区面积20 m2, 长5 m, 宽4 m, 行距50 cm, 株距40 cm, 密度5 株· m-2。四周设保护行。
1.3.2 田间管理 在苗期进行匀苗工作, 2-3片真叶期间苗, 每窝留2~3株, 5叶期定苗1株。中耕一次, 培土一次, 结合中耕培土追肥一次, 1 hm2土地施尿素300 kg。5叶期防治菜粉蝶一次。
2013年9月至2014年5月, 选取具有品种特性的10株芜菁甘蓝随机调查形态学特征。子叶长、子叶宽、莲座期株高、莲座期株幅、小裂叶对数、莲座期叶数、叶长、叶宽、块根长、块根宽、单根重、单株重、单产、开花期株高、有效分枝位、一次分枝数、二次分枝数、主花序长、主花序角果数、单株角果数、角果长度、角果宽度、单角果种子数、种子产量、种子千粒重、播种-块根收获期、播种-种子成熟期27个数量性状(数值型性状)采用测量法; 子叶颜色、下胚轴颜色、侧芽大小、叶型、叶形、叶顶端形状、叶缘齿状、叶缘波纹、叶裂刻、叶裂回数、叶面、叶面刺毛、叶面蜡粉、叶面光泽、叶簇、株型、分蘖、叶色、叶柄色、叶柄切面、叶瘤大小、抽薹性、花茎颜色、花色、角果颜色、角果落粒性、种皮颜色、商品熟性、根形、根肉色、根皮色、须根、块根口感、块根质地34个质量性状按照《叶用和薹(籽)用芥菜种质资源描述规范和数据标准》[6]进行性状观察记载赋值, 按各性状值代码进行数字化再进行分析。
数量性状采用Excel 2007和SPSS v10.0软件统计出其最大值、最小值、平均值、标准差、极差和变异系数; 质量性状利用NTSYS-pc v2.2软件进行聚类分析。
分析28份芜菁甘蓝种质资源的27个数量性状发现, 不同材料间存在很大差异, 同时不同性状在不同材料间表现出的多样性也不一致(表2)。
| 表2 芜菁甘蓝种质资源中27个数量性状的遗传多样性评价 Table 2 Assessment on genetic diversity in 27 quantitative traits of Brassica napobrassia germplasm resources |
不同材料间的变异系数存在很大差异。单株角果数的变异程度最大, 达到64.1%, 变幅为154.0~4 292.0粒; 其次为小裂叶对数, 其变异系数为57%, 变幅为0~16.0对; 角果宽度的变异系数为51.7%, 变幅为0.18~2.50 cm; 子叶长的变异系数为48.7%, 变幅为0.95~7.50 mm; 子叶宽的变异系数为46.9%, 变幅为1.50~10.0 mm。以上5个数量性状的变异系数均在45%以上, 变幅范围较大。
有效分枝位的变异系数为5.6%, 变幅为1.5~36.5 cm; 播种-种子成熟期的变异系数为3.5%, 变幅为250~271 d; 播种-块根收获期的变异系数为3.1%, 变幅为86~115 d; 单株重的变异系数为2.9%, 变幅为0.5~6.5 kg; 单根重的变异系数最小为2.4%, 变幅为0.2~4.5 kg。以上5个数量性状的变异系数均< 10%, 变幅范围较小。剩余17个数量性状的变异系数均在10%~40%。分析27个数量性状统计变量, 如平均值、最大值、最小值、标准差、极差和变异系数可见, 28份芜菁甘蓝种质资源的遗传多样性较为多样, 遗传改良利用潜力强健, 可以为提高芜青甘蓝的品质和产量提供实际意义。
对28个芜菁甘蓝材料的27个数量性状进行赋值, 并利用UPGMA法对供试材料进行聚类分析(图1), 可将28份参试材料聚为4类。第Ⅰ 类具有子叶为绿色、侧芽小、叶型为板叶、叶顶端形状都为圆形、根形椭圆及须根少等特点, 由贵州省草业研究所的威宁芜菁甘蓝(1号)和花溪芜菁甘蓝(3号), 百绿集团的拿破仑灰萝卜(4号), 重庆的江口洋萝卜(7号), 道真的洛龙洋萝卜(8号)、河口洋萝卜(10号)和三桥灰萝卜(11号), 青海省畜牧兽医科学院的青海玉树芜菁(14号), 新疆的卡马古芜菁(15号), 内蒙古紫皮蔓菁(16号), 昭通的昭通洋萝卜(18号), 清镇的新店灰萝卜(19号), 新西兰的KRC-6004(25号)和KRC-6063(26号)共14份参试材料组成。
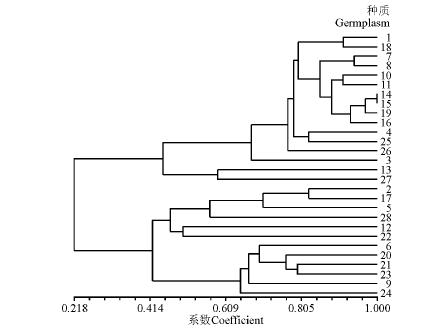 | 图1 28个芜菁甘蓝种质聚类Fig.1 Dendrogram of the 28 germplasm resources using a clustering analysis according to morphological traits |
第Ⅱ 类是由来自邢台兴达种业有限公司的京丰1号(13号)和新西兰的KRC-7860(27号)组成, 具有叶面平滑、叶面无刺毛且少蜡粉、叶面有光泽及叶簇呈半直立等特点。
来自凉山彝族自治州畜牧兽医科学研究所的四川园根(2号)、百绿集团的德国园根(5号)、温州市神鹿种业有限公司的温州盘菜(12号)、凉山的凉山芜菁(17号)、重庆的秀山大梗菜(22号)、新西兰的KRC-7809(28号)共6份参试材料聚为第Ⅲ 类, 此类具有分蘖能力中等、叶色主要呈黄绿色、角果颜色为浅绿色、根肉色为白色及须根少等特点。
第Ⅳ 是由来自重庆的武陵大头菜(6号)、道真的阳溪芥菜(9号)、重庆的清溪黄心芥(20号)、松桃的瓦溪大头菜(21号)、独山的独山大头菜(23号)及新西兰
的KRC-3960(24号)共6份材料组成, 具有叶缘有复锯齿和大波纹、叶裂刻呈全裂或深裂、叶裂回数均为一回及有叶瘤、种皮颜色为红褐色等特点。
种质资源遗传多样性是作物遗传育种的物质基础, 不仅有助于种质资源的管理、评价和利用, 更有利于优异种质的创新研究[7, 8, 9, 10, 11]。在小麦[12, 13](Triticum aestivum)、苜蓿[14-15](Medicago)、斑茅[16](Erianthus arundinaceum)、大豆[17-18](Glycine max)和燕麦[19-20](Avena sativa)等植物上, 均有应用数量性状探讨种质遗传多样性的报道。本研究对国内外育成品种28份芜菁甘蓝种质资源27个数量性状进行遗传多样性研究, 发现芜菁甘蓝种质资源在不同品种的不同性状上表现出不同程度的多样性, 可以作为芜菁甘蓝种质资源数量性状分析的主要性状指标。各性状中单株角果数的变异系数最大, 其次是小裂叶对数及角果宽度, 说明这3个性状的变异性大, 遗传性丰富, 蕴藏着较大的选择空间, 对种质变异和创新的贡献率越大, 可供选择和利用的潜力大, 在常规育种中, 可以把变异系数较大的性状作为其数量分析的主要性状, 通过对28份芜菁甘蓝数量性状的分析, 能选育出期望得到的变异类型的概率较大。聚类分析结果表明, 28份供试材料可划归为四大类, 具有同一性状的大部分材料聚在同一类。类内材料具有相同或相似的形态性状, 而类间的形态性状差异较明显。
The authors have declared that no competing interests exist.
| [1] |
|
| [2] |
|
| [3] |
|
| [4] |
|
| [5] |
|
| [6] |
|
| [7] |
|
| [8] |
|
| [9] |
|
| [10] |
|
| [11] |
|
| [12] |
|
| [13] |
|
| [14] |
|
| [16] |
|
| [17] |
|
| [18] |
|
| [19] |
|
| [20] |
|