

{kind=link}
{kind=link}
{kind=link}
放牧对天山北坡山地草原生态系统土壤δ15N的影响
[张青青1  , 安沙舟
, 安沙舟1 , 于辉1 , 吐尔逊娜依·热依木1 , 张沛2 , 张玲卫1 , 禹朴家3 ]
, 安沙舟, 于辉|
|


第一作者:张青青(1983-),女,新疆和静人,副教授,博士,主要从事干旱区生态恢复与保护的研究。E-mail:[email protected]
测定了天山北坡山地草原优势植物种针茅( Stipa capillata)、羊茅( Festuca ovina)、短柱苔草( Carex turkestanica)、博乐绢蒿( Seriphidium borotalense)根际0-10 cm土壤和非根际10-20、20-30 cm土层土壤稳定性N同位素和全氮含量,通过对比放牧和围封草地优势植物根际土壤δ15N的变化,分析不同利用方式对不同植物根际土壤和不同土层深度土壤δ15N的影响。结果表明,天山北坡山地草原生态系统土壤δ15N值在2.25‰~16.03‰变化,放牧显著降低了表层0-10 cm土壤δ15N值;放牧草地0-30 cm土壤δ15N值(7.14‰±0.67‰)整体上比围封(6.95‰±0.34‰)增加了2.8%,但差异不显著( P>0.05)。放牧和围封条件下天山北坡山地草原土壤δ15N值均随着土层深度的加深而显著增加( P<0.05),且土壤δ15N值与土壤全氮含量呈显著的负相关( P<0.05)。放牧对天山北坡山地草原不同植物种根际土壤δ15N的影响程度不同,放牧草地的针茅和羊茅0-30 cm土壤δ15N分别比围封草地的高17%和53%,但放牧降低了短柱苔草和博乐绢蒿0-30 cm土壤δ15N,且对短柱苔草的降低作用显著。
, An Sha-zhou, Yu HuiThe total N and δ15N of rhizosphere soil 0-10 and 10-20, 20-30 cm of Stipa capillata, Festuca ovina, Carex turkestanica and Seriphidium borotalense in the mountainous grassland ecosystems over the northern Tianshan Mountains were measured. δ15N of dominant plants' rhizosphere soil in grazing and fencing grassland was compared to reveal the impacts of different usage patterns on δ15N of rhizosphere soil in different plants and different soil layer. The results indicated that the range of soil δ15N of mountainous grassland ecosystems over the northern Tianshan Mountains were from 2.25‰ to 16.03‰. Grazing significantly decreased δ15N in 0-10 cm soil. The δ15N in 0-30 cm soil of grazing grassland (7.14‰±0.67‰) increased by 2.8% than that of fencing grassland (6.95‰±0.34‰) although there was no significant difference between them. The δ15N in soil significantly increased with the increasing of soil depth in both grazing and fencing grassland and it had significantly negative correlation with soil total N. The impacts of grazing on δ15N in rhizosphere soil of different plants were different. The δ15N in the 0-30 cm rhizosphere soils of Stipa capillata and Festuca ovina in the grazing grassland were 17% and 53% higher than that in fenced grassland respectively. However, the δ15N in the 0-30 cm rhizosphere soils of Carex turkestanica and Seriphidium borotalense were lower than that in fenced grassland. The measurement of δ15N supported an aggregative indicator of N cyclic process in a typical area for the ecosystem and showed that grazing changed the N cycle of grassland ecosystem.
物质循环是生态系统的基本功能, 氮(N)是物质的基本组成元素。地球表面氮素的生物地球化学循环过程及其环境效应是当前全球变化中区域尺度上研究的重要内容, 国内外许多核心项目也都把氮素循环作为主要研究内容。氮是植物生长发育最重要的元素之一, 在自然生态系统中, 大部分的氮只有首先被矿化才能被生物所利用。草地生态系统可利用氮素的供应与草地生态系统的生产力有直接相关性, 在很大程度上是第一性生产力的重要限制因子[1]。植物和土壤中15N自然丰度(δ 15N)是探明生态系统N循环与评价生态系统氮效应的一个重要指标, δ 15N在N的输入、转化和输出过程中由于同位素的分馏而引起δ 15N值的差异[2]。例如, 动物的践踏、啃食和富N粪尿的增加等都会直接或间接影响土壤的N循环, 从而会不同程度地造成δ 15N值的变化。而放牧是草地生态系统最主要的土地利用方式之一, 对草原生态系统N循环过程的改变起着重要的作用, 为理解生态系统中的N循环提供有用的信息[3, 4, 5, 6]。近年来, 国内外进行了大量针对草地生态系统中氮素循环的研究[6, 7, 8, 9], 包括草地生态系统氮素循环的通量和过程的系统研究[10, 11]以及氮肥草地生态系统结构与功能的调控机理方面的研究[12, 13]等, 而且也已经有不少学者开展了放牧对草地生态系统N素分配的影响[10, 14, 15]。从全球范围来看, 影响δ 15N格局的因素主要有气候、时间、地形和土地利用等[16, 17, 18, 19]。而在区域范围内, 土壤δ 15N值主要受土壤N输入(包括牲畜粪尿、氮沉降和生物固氮)、输出、转化中的分馏作用以及植物的生长生理等一系列生物地球化学循环的影响[2, 20]。
新疆作为我国五大牧区之一, 天然草地分布区域广, 总面积5.725 88× 107 hm2, 占全国草地总面积的14.6%[21, 22]。由于受人类利用方式和自然因素的影响, 草地生态系统的承载力常常被忽略, 使该区大部分天然草地长期处于不同程度的过度放牧状态, 导致草地生态系统退化。草地生态系统服务功能的降低常常是由于其自身的退化所引起的, N素循环也常常由于生态系统处于开放状态而造成N素流失, 从而使生态系统结构和功能的耦合机制失调。种类成分(如不同功能型植物之间的替代)以及植物生长特性的变化进一步造成N素循环过程的变化。因此, 有必要选取典型草原生态系统通过成对比较放牧和围封条件下山地草原优势植物种根际土壤δ 15N的变化, 分析不同草地利用方式对不同植物根际土壤δ 15N的影响是否存在差异, 分析放牧对草原生态系统N循环过程的影响, 为提高草地生产力、合理并持续利用草地资源提供理论依据。
研究区位于天山北坡乌鲁木齐县甘沟乡的山地草原生态系统(43° 33'56″ N, 87° 13'10″ E), 海拔1 524 m, 位于亚欧大陆腹地, 远离海洋, 显著的大陆性气候, 其特点是冬季寒冷漫长, 夏季炎热, 气温年、日较差大, 春秋季气温变化剧烈, 光照充足, 热量丰富, 年降水量200~550 mm。
试验样地选择自由放牧和围封保护的山地草原为研究对象。研究区地带性土壤为淡栗钙土, 土壤含水量为4.99%~7.95%, pH为8.05~8.52, 有机质11.823~23.061 g· kg-1, 全氮0.563~1.298 g· kg-1, 全磷0.690~0.802 g· kg-1。草地建群植物有针茅(Stipa capillata)、羊茅(Festuca ovina)、短柱苔草(Carex turkestanica)、博乐绢蒿(Seriphidium borotalense), 且所选择样地无固氮植物出现。采样区于20世纪90年代建立围栏, 分别进行自由放牧和围栏封育恢复试验。试验前后, 自由放牧和围栏封育区没有添加化肥, 土壤性质也没有显著差异。自由放牧样地和围栏封育样地相邻, 放牧在每年的5-6月和9-11月进行。
| 表1 放牧与围栏样地土壤理化性质 Table 1 Physical and chemical properties of the soil in grazing and fencing plot |
在一处地势较为平坦的区域, 选取3个样地, 每个样地4个1 m× 1 m的样方, 于2013年8月24日, 用土钻分别取每个样方中针茅、羊茅、短柱苔草、博乐绢蒿4种植物根际0-10 cm土壤和非根际10-20、20-30 cm土层土壤, 每种植物根际周围土壤取3钻混合, 作为一个重复。将其放入封口袋后带回实验室。在实验室内自然风干后, 研磨, 再过孔径0.18 mm的土壤筛, 然后取筛好的土壤样品5 g放在封口袋中保存, 作为土壤稳定性N同位素待测样品[8, 10]。另外, 将4种植物根际周围的土壤分别取少量混合, 与土壤稳定性N同位素样品一样分层处理, 作为土壤全氮的待测样品。
将适量过筛处理后的土壤稳定性N同位素样品(取样量根据样品的氮含量而定)装入专用的金属锡舟中, 然后在配有自动进样装置的EA1112HT-MAT253高温裂解元素分析仪和稳定性同位素质谱仪(Thermo Scientific, USA)上进行测试, 标样连续测试精度小于0.3‰ [8, 10]。EA反应炉温度为960 ℃, GC柱温为50 ℃, 载体流速为85 mL· min-1。测得的δ 15N值是一个相对比例, 用公示表达为:
δ 15N=(Rsample/Rstandard-1)× 1 000‰ .
其中:Rsample、Rstandard分别表示样品和标准物(空气)之间15N/14N的比值。
用SPSS 13.0软件进行单因素方差分析(One-way ANOVA)和最小显著差异法(LSD)比较检验放牧与围封、不同土层深度、不同植物之间土壤δ 15N值的差异程度, 用Pearson相关系数分析土壤δ 15N值和土壤全氮含量的相关关系。
天山北坡山地草原生态系统, 土壤δ 15N值的变化范围在2.25‰ ~16.03‰ , 围封样地土壤δ 15N值是(6.95‰ ± 0.34‰ ), 放牧样地土壤δ 15N值是(7.14‰ ± 0.67‰ ), 放牧后0-30 cm土壤δ 15N值整体上比围封增加了2.8%, 但无显著差异(P> 0.05)。
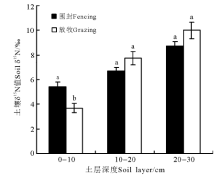
放牧对不同深度土壤δ 15N的影响有所差异(图1)。无论是放牧样地还是围封样地, 天山北坡山地草原生态系统土壤δ 15N值随着土层深度的增加而不断增加。放牧后, 10-20和20-30 cm土壤δ 15N均比围封样地土壤δ 15N值有所增加, 均增加15%, 但差异不显著(P> 0.05)。而0-10cm土壤δ 15N值在放牧后有所降低, 比围封样地土壤δ 15N值低32%, 且差异显著(F=6.946, P< 0.05)。
 | 图1 山地草原生态系统不同深度土壤δ 15N的变化注:不同小写字母表示同一土层不同处理间差异显著(P< 0.05)。Fig. 1 Soil δ 15N of different soil layer in mountainous grassland ecosystemsNote: Different lower case letters in the same soil layer indicate significant difference between fencing and grazing at 0.05 level. |
放牧对不同植物种根际土壤δ 15N的影响程度不同(图2)。围封样地, 针茅和羊茅根际0-30 cm深度土壤δ 15N值分别为8.19‰ 和6.22‰ , 比放牧样地土壤δ 15N值增加了17%和53%, 且针茅根际0-30 cm深度土壤δ 15N值在放牧和围封样地差异显著(F=16.607, P< 0.05), 羊茅差异极显著(F=28.506, P< 0.01)。而短柱苔草和博乐绢蒿根际0-30 cm土壤δ 15N值在围封样地分别为6.47‰ 和6.91‰ , 放牧后样地0-30 cm土壤δ 15N值均相比围封样地有所降低(图2), 分别降低了28%和30%, 且短柱苔草根际0-30 cm深度土壤δ 15N值在放牧和围封样地差异极显著(F=27.930, P< 0.01), 博乐绢蒿差异不显著(P> 0.05)。
 | 图2 不同植物不同土层根际土壤δ 15N的变化注:不同大、小写字母分别表示同一土层不同处理间差异极显著(P< 0.01)和显著(P< 0.05)。Fig.2 Soil δ 15N of different plant rhizosphere(different soil layer)Note: Different capital and lower case letters in the same soil layer indicate significant difference between fencing and grazing at 0.01 and 0.05 level, respectively. |
从放牧与围封对不同植物种根际不同深度土壤δ 15N的影响来看(图2), 短柱苔草和博乐绢蒿根际0-10 cm土壤和非根际10-20、20-30 cm土壤的δ 15N值在放牧条件下均比围封有所降低, 其中短柱苔草根际0-10 cm土壤的δ 15N值在放牧条件比围封降低了43%, 差异显著(F=8.447, P< 0.05), 博乐绢蒿根际0-10 cm和非根际10-20 cm土壤的δ 15N值在放牧条件下比围封分别降低了46%和38%, 差异显著(分别为F=14.627, P< 0.05; F=15.554, P< 0.05), 其余深度的土壤δ 15N值在放牧和围栏条件下差异不显著(P> 0.05)。针茅和羊茅根际0-10 cm土壤的δ 15N值在放牧条件下均比围封有所降低, 分别降低19%和14%, 但差异不显著; 而羊茅非根际10-20和20-30 cm土壤的δ 15N值在放牧条件下比围封有所增加, 增加幅度分别为75%和74%, 差异显著(分别为F=32.584, P< 0.01; F=12.908, P< 0.05); 针茅非根际10-20和20-30 cm土壤的δ 15N值在放牧条件下比围封有所增加, 增加幅度分别为48%和16%, 但10-20 cm土壤的δ 15N值在放牧和围封条件下差异不显著, 20-30 cm差异显著(F=8.505, P< 0.05)。
把所有放牧与围封条件下不同植物根际不同深度的土壤δ 15N值放在一起考虑, 土壤δ 15N值与全氮值的Pearson相关系数为-0.633, 存在显著的相关关系(R2=0.400 1, P< 0.01)。无论在放牧还是围封条件下, 随着土壤全氮含量的增加土壤δ 15N值均表现为先降低后增加, 且放牧条件下的多项式拟合(y=16.597x2-31.476x+17.543, R2=0.771 2)比围封条件下的多项式(y=23.508x2-34.06x+17.764, R2=0.394 5)拟合程度高(图3)。
 | 图3 土壤全氮含量与土壤δ 15N的相关关系Fig.3 Relationship between soil total nitrogen content and soil δ 15N |
放牧是草地生态系统最主要的土地利用方式之一, 对草原生态系统N循环过程的改变起着重要的调节作用。放牧提高土壤的净氮矿化速率, 促进淋溶、挥发和反硝化等过程, 增加贫化15N的流失, 形成一个开放的氮系统, 从而引起土壤δ 15N值的增加[16, 23, 24]。但也有研究表明, 长期的放牧导致氮循环速率降低, 土壤的δ 15N值也随之降低[25, 26]。本研究表明, 天山北坡山地草原生态系统0-30 cm土壤δ 15N值在放牧条件下整体上比在围封条件下有所增加, 而表层0-10 cm土壤δ 15N值在放牧条件下比围栏降低了32%, 且差异显著(F=6.946, P< 0.05)。本研究所选择的天山北坡山地草原生态系统的放牧样地, 仅有极少量的牲畜粪尿, 有研究表明, 较少的或呈斑块化分布的排泄物对生态系统土壤δ 15N值的作用相对较小[27]。且选择的放牧样地与围封样地是相邻样地, 因此氮沉降所造成的差别也可以忽略不计。由于所选择的样地没有固氮植物出现, 因此生物固氮的影响也可以忽略不计。另外, 仅表层0-10 cm土壤δ 15N值在放牧条件下比围封降低, 而10-20和20-30 cm土壤δ 15N值在放牧条件下比围封有所增加(图1)。且不管是在放牧条件下还是围封条件下, 土壤δ 15N值均随着土层深度的加深而显著增加, 这与国内外的许多研究[28, 29]结果相一致。在围封条件下, 10-20 cm土壤δ 15N值比0-10 cm增加了24%, 20-30 cm土壤δ 15N值比0-10 cm增加了62%, 而在放牧条件下, 10-20 cm土壤δ 15N值比0-10 cm增加了110%, 20-30 cm土壤δ 15N值比0-10 cm增加了172%, 土壤δ 15N值随着土层深度的加深而增加的速度比在围封条件下快。综上可知, 放牧对草原生态系统N循环过程的改变有着重要的调节作用, 降低土壤δ 15N值。
短柱苔草根际0-10 cm土壤、博乐绢蒿根际0-10 cm和非根际10-20 cm土壤的δ 15N值在放牧条件下均比围封有所降低, 且差异显著。针茅和羊茅根际0-10 cm土壤的δ 15N值在放牧条件下也比围封有所降低, 但差异不显著。不同植物会不同程度地影响土壤δ 15N值的变化, 但从总体上来看, 不同植物根际的表层0-10 cm土壤, 均在放牧条件比围封有所降低, 也就是说放牧对不同植物根际土壤表层0-10 cm土壤δ 15N值的降低作用是一致的。干旱区土壤水分含量有限, 土壤微生物活动相对较弱, 因此, 土壤中的N主要来自植物残体的贡献。本研究结果与15N贫化的凋落物输入导致表层土壤较低的15N含量的结果也恰好相符。另外, 放牧与围栏条件下, 土壤全氮含量均在表层0-10 cm土壤中最高(放牧条件下土壤全氮量为1.298 g· kg-1; 围栏条件下, 土壤全氮量为0.711 g· kg-1), 而相应的放牧与围栏条件下0-10 cm土壤δ 15N是各个土层最低的, 分别为5.36‰ 和5.40‰ , 这恰好与土壤δ 15N值和全氮值存在显著的负相关关系相一致, 土壤全氮含量越高, 土壤δ 15N值越低。这与吴田乡和黄建辉[26]的观点一致, 认为放牧后0-10 cm土壤δ 15N值的显著降低可能与长期放牧有关, 长期放牧引起N循环速率减小, 从而使土壤δ 15N值降低。
本研究以山地草原生态系统为研究对象, 通过对比放牧和围封条件下山地草原4种优势植物种土壤δ 15N的变化, 分析不同利用方式对不同植物土壤δ 15N的影响, 探讨草原生态系统土壤δ 15N值在放牧与围栏条件存在差异的原因, 得到以下结论:天山北坡山地草原生态系统土壤δ 15N值在2.25‰ ~16.03‰ 变化, 放牧显著降低了表层0-10 cm土壤δ 15N值, 0-30 cm土壤δ 15N值(7.14‰ )整体上比围封(6.95‰ )增加了2.8%, 但差异不显著, 不管在放牧还是围栏条件下, 土壤δ 15N值均随着土层深度的加深而显著增加, 且土壤δ 15N值与土壤全氮量显著负相关。放牧对不同植物种根际土壤δ 15N的影响程度不同, 放牧后, 针茅和羊茅根际0-30 cm土壤δ 15N值分别比围封增加17%和53%, 但放牧降低了短柱苔草和博乐绢蒿0-30 cm土壤δ 15N值, 且对短柱苔草的降低作用显著。δ 15N值的测定为生态系统提供了一个区域范围内氮循环过程的综合指标, 反映出放牧改变了草原生态系统的N循环速率。
The authors have declared that no competing interests exist.
| [1] |
|
| [2] |
|
| [3] |
|
| [4] |
|
| [5] |
|
| [6] |
|
| [7] |
|
| [8] |
|
| [9] |
|
| [10] |
|
| [11] |
|
| [12] |
|
| [13] |
|
| [14] |
|
| [15] |
|
| [16] |
|
| [17] |
|
| [18] |
|
| [19] |
|
| [20] |
|
| [21] |
|
| [22] |
|
| [23] |
|
| [24] |
|
| [25] |
|
| [26] |
|
| [27] |
|
| [28] |
|
| [29] |
|