{kind=link}
季节性放牧对草地植物多样性与功能群特征的影响
[刘玉1  , 刘振恒
, 刘振恒2 , 邓蕾1 , 武高林1 ]
, 刘振恒]
|
|


第一作者:刘玉(1988-),男,陕西洛南人,在读博士生,主要从事草地植被生态与水土保持方面的研究。E-mail:[email protected]
针对放牧对高寒草地生态系统的影响,通过比较暖季放牧(6月-10月)和冷季放牧(11月-翌年5月)下典型高寒草地物种多样性和功能群特征的变化,分析了高寒草地植物群落对季节性放牧的响应机制。结果表明,暖季放牧草地中物种多样性指数为1.92,均匀度指数为0.78,明显高于冷季放牧草地的1.81和0.74,而冷季放牧草地的丰富度指数为17.45,高于暖季放牧草地的16.45。冷季放牧草地的地上生物量为246.61 g·m-2,莎草类和豆科类功能群的生物量比例分别为66.85%和15.86%,其值均高于暖季放牧草地。暖季放牧草地植物总密度为2 064株·m-2,明显高于冷季放牧草地的1 394株·m-2。合理的放牧强度下,暖季放牧相比冷季放牧有利于草地群落物种多样性和均匀度的维持,但冷季放牧草地有利于草地群落地上生物量的积累。
To determine the effects of grazing on alpine grassland ecosystem, the response of the diversity and the functional group to different seasonal grazing which was divided warm-season grazing (June to October) and cold-season grazing (the other months) in alpine grassland was analyzed. The results showed that the diversity and evenness index in warm-season grassland was significantly greater than that in cold-season grassland, however, the richness index was reverse. showed a contrary trend. Above-ground biomass, the proportion of sedge and legume function group in warm-season grazing grassland was significantly greater than that in cold-season grazing grassland. Our results suggest that under a suitable grazing intensity, warm-season grazing would be beneficial to maintain the higher species diversity and evenness sustainability, and cold-season grazing would be beneficial to accumulate above-ground biomass in alpine grassland.
草地作为畜牧业生产的主要物质基础, 同时也是重要的生态屏障, 对维持区域生态系统平衡起着重要作用[1]。随着我国人口的不断增加, 居民膳食结构也发生了巨大变化, 其显著特征是粮食等主食消费减少, 肉、蛋、奶等动物性食品消耗增加[2]。对畜产品需求的日益增加, 使得草地生态系统面临着越来越大的放牧压力, 很多草地都已经出现过度放牧、严重恶化的现象[3]。如何转变牧区传统的生产方式, 促进草业发展, 发挥其潜在的生态和生产功能, 是当前一个重要而紧迫的课题[2]。
放牧是人类利用高寒草地的主要方式之一, 是草原生物生产转化为经济效益无可替代的过程, 但过度的放牧利用会导致草地的退化[4]。放牧引起草地的退化是一个渐变过程, 会经过一系列的演替阶段, 主要是由于家畜的过度采食和践踏引起草地生态系统动植物及微生物组分的改变[5, 6]、土壤结构改变和养分失衡[1, 7]。过度放牧导致草地生态系统结构和功能的严重丧失, 对区域经济发展乃至整个生态系统都会产生深刻影响[8]。近年来, 针对放牧草地生态系统已有大量研究, 放牧强度[6, 9, 10, 11]、放牧制度[8]和放牧时间[3, 12]均对草地群落有直接影响。放牧强度的增加导致群落地上部分生物量和净生产力显著降低, 然而适当放牧可以增加草地的地上净生产力[10]。多数研究集中于放牧强度(梯度)对草地的影响, 而关于季节性放牧对草地多样性及功能群特征的影响缺少相对系统的研究。
高寒草地由于特殊的地理气候条件, 生态系统脆弱, 抗干扰能力差, 植被一旦遭受破坏, 靠其自然恢复极为困难[13, 14]。季节性放牧是青藏高原高寒地区畜牧业生产的主要方式之一。不同季节中, 放牧家畜对植物的采食策略及模式不同, 增加了植物群落微生境的异质性, 从而使得草地结构、功能等发生改变。本研究针对青藏高原地区玛曲县的高寒草地放牧生态系统, 通过分析比较暖季和冷季放牧利用方式, 探讨季节性放牧对高寒草地植物群落的影响, 以期揭示高寒草地群落物种多样性和功能群特征对季节性放牧的响应机制, 为高寒草地生态系统科学放牧制度的建立及草地的合理利用提供理论指导。
研究区域位于青藏高原东缘的甘肃省甘南州玛曲县境内, 地理位置33° 38' N、102° 06' E。该地区海拔约3 500 m, 属于大陆性季风气候, 年平均温度约为2.2 ℃, 最高温度在7月份(均温11.7 ℃), 最低温度在1月份(均温-10 ℃), 年平均降水量约672 mm, 主要集中在6-8月, 属于高寒湿润区。全年日照时长约为2 580 h, 年平均霜日大于270 d, 无绝对无霜期。土壤类型为高寒草甸土, 植被组成主要是以禾本科的羊茅属(Festuca)、早熟禾属(Poa)和莎草科的嵩草属(Kobresia)为优势种, 豆科苜蓿属(Medicago)、野豌豆属(Vicia)和菊科的风毛菊属(Saussurea)为伴生种, 同时伴生有大量其它杂类草。
试验样地选择研究区内典型的高寒草地, 选择一处长势均匀的天然草地设置为两个处理, 暖季放牧和冷季放牧, 每个处理各设置4个100 m× 100 m的围栏样地进行试验。放牧家畜为藏羊, 放牧时间为暖季放牧(6月-10月)和冷季放牧(11月-翌年5月), 期间进行连续放牧, 其余时间样地均处于围栏休牧状态[11]。放牧强度为每公顷草地上暖季放牧2~2.5羊单位, 冷季放牧1~1.5羊单位, 根据前期预先的测量计算并参考相关文献确定草地利用率约为30%[15]。
在两种处理样地中各进行完一次冷季放牧和暖季放牧处理后, 于2013年10月暖季放牧结束时, 对两个处理的8个样地进行群落调查。在每个样地内随机选择一个60 m× 80 m的调查小区, 在小区的对角线上设置5个0.5 m× 0.5 m 的样方进行群落调查, 各样方间隔为10 m。测定各放牧样地内植被群落的物种种类、多度、盖度、高度, 将植物分种齐地面刈割, 然后将样品带回实验室, 于80 ℃下烘至恒重后称量, 记录为地上生物量[10]。试验中将群落分为禾草类、莎草类、豆科类和杂类草4个功能群[5], 并根据烘干后称量的各物种地上生物量计算各功能群的生物量比例。
变异系数=标准差/平均值× 100%;
多样性[16]计算:
Patrick丰富度指数(R1):R1=S;
Margalef丰富度指数(R2):R2=(S-1)/ln N;
Menhinick丰富度指数(R3):R3=S/N;
Shannon-Wiener多样性指数(H1):
H1=-∑ (Pi)lnPi;
Simpson多样性指数(H2):
H2=∑ [Ni(Ni-1)/N(N-1)];
Pielou均匀度指数(E1):E1=H1/ln S;
Alatalo均匀度指数(E2):
E2 = (N2-1) / (N1-1)。
式中:S为每个样方中的物种数, N为样方中所有物种的总个体数, Ni为样方中的物种i个体数, Pi为样方中第i种个体数占全部物种个体数N的比例。
用Excel 2010统计两种放牧草地中各功能群的物种数、密度、生物量及其比例, 用SPSS 18.0进行独立样本t检验比较不同放牧草地群落物种多样性、丰富度、均匀度、生物量及密度的差异显著性(P< 0.05)。
从暖季和冷季放牧草地群落的3个丰富度指数变化来看, 冷季放牧草地的物种丰富度指数均高于暖季放牧草地的(表1)。3个丰富度指数的变异系数变化也表现为冷季放牧草地的变异程度大于暖季放牧草地的。相比3个物种丰富度指数的变化, 两个多样性指数的变化呈现暖季放牧草地大于冷季放牧草地, 且Simpson多样性指数在暖季放牧草地(0.78)显著高于冷季放牧草地的(0.74)(P< 0.05)。从两个多样性指数的变异系数来看, Shannon-Wiener多样性指数的变异系数在暖季草地中的变异程度大于冷季放牧草地, 而Simpson多样性指数的变化则相反。两种均匀度指数变化和多样性指数相似, 均匀度指数均显示为暖季放牧草地的显著高于冷季放牧草地的(P< 0.05)。Pielou均匀度指数在暖季放牧草地中的变异程度大于冷季放牧草地, 但Alatalo均匀度指数的变异系数则相反, 呈现冷季放牧草地大于暖季放牧草地。
| 表1 不同季节放牧草地物种多样性变化 Table 1 The changes of diversity in different seasonal grazing grassland |
冷季放牧草地群落的总生物量显著高于暖季放牧草地的(P< 0.05)。禾草、莎草、豆科和杂类草功能群的生物量均表现出冷季放牧草地大于暖季放牧草地。其中, 莎草功能群生物量在冷季放牧草地为66.85 g· m-2, 显著大于暖季放牧草地的30.63 g· m-2(P< 0.05)。禾草、豆科和总生物量的变异系数表现为暖季放牧草地大于冷季放牧草地, 而莎草和杂类草功能群的变异程度则表现为冷季放牧草地大于暖季放牧草地。
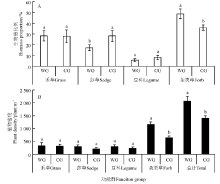
不同功能群的生物量比例可以看出(图1A), 暖季放牧草地中禾草功能群的生物量比例(28.27%)略高于冷季放牧草地的(27.84%)。同时, 暖季放牧草地中杂类草功能群的生物量比例显著高于冷季放牧草地的(P< 0.05)。莎草功能群和豆科功能群生物量比例则是冷季放牧草地大于暖季放牧草地。其中, 莎草功能群生物量比例在冷季放牧草地中达到28.16%, 显著高于暖季放牧草地中的(17.25%)(P< 0.05)。
 | 图1 不同季节放牧草地群落的功能群特征变化注:同一功能群中的不同小写字母表示两种放牧草地间差异显著(P< 0.05)。Fig.1 Characteristics of functional groups in different seasonal grazing grasslandsNote: Different lower case letters for same functional group indicate significant difference between two grazing grasslands at 0.05 level. |
暖季放牧草地群落中植物总密度达到了2 064株· m-2, 显著高于冷季放牧草地的1 394株· m-2(P< 0.05)。禾草、莎草、豆科、杂类草功能群的植物密度均表现为暖季放牧草地大于冷季放牧草地, 其中杂类草功能群密度在暖季放牧草地中为1 157株· m-2, 显著高于冷季放牧草地的642株· m-2(P< 0.05)。
放牧作为高寒草地的主要干扰因素, 可通过调节群落的物种多样性和功能多样性[17, 18]影响资源的转换和分布, 从而影响草地群落结构和物种组成[19, 20]。根据生态学中关于植物对家畜采食响应的研究, 在植物生长季中, 家畜采食的时间不同, 植物的生长反应不同; 且同一时间, 不同植物的反应也不相同[11]。本研究表明, 不同放牧季节的高寒草地, 群落的多样性出现不同的变化模式, 暖季放牧草地的物种多样性高于冷季放牧草地的(表1)。在暖季放牧草地中家畜的牧食行为抑制了草地优势种(禾草和莎草功能群)的生长, 打破了草地群落组分种间的竞争地位, 为群落中非优势种的生长提供了机会[11]。另外也有研究表明, 放牧也通过影响植物的有性繁殖来影响群落组成, 如在羊草(Leymus chinensis)草地, 放牧导致植物的种子生物量、结实数和生殖枝的分化率下降, 草地的地下生物量分配比例增加, 而且同化器官、非同化器官和种子生物量比例下降, 从而影响草地的自然更新[21]; 放牧草地中植物的有性繁殖活动, 如物候期、花的形态及数目、结籽率等均会受到不利影响[22]。暖季放牧草地中, 由于家畜采食, 很多植物的有性生殖比例减小。冷季放牧草地中, 大多数物种的种子都能成熟, 家畜在采食过程也可能携带并转移一些植物种子, 使一些新物种可以成功入侵, 从而表现为冷季放牧草地的丰富度指数高于暖季放牧草地的。
功能群通常被认为是与生态系统的某种功能相关的一类特定的物种群, 就草地物种组成而言, 功能群决定着生态系统的生产力和稳定性[23]。本研究中的放牧草地主要以耐牧性较强的禾草功能群和莎草功能群为主, 其特点是根茎繁殖能力强、耐践踏、再生能力强。因此, 暖季放牧草地中这些优势植物依然能保持良好生长。但莎草类功能群的生物量比例在冷季放牧草地显著高于暖季放牧草地的, 可以看出, 家畜在暖季放牧草地中采食了大量的莎草类植物, 造成莎草类功能群生物量比例显著减少。也有研究表明, 季节性放牧对草地群落的影响与家畜的嗜食性有关[24]。放牧条件下, 适口性好的禾草和莎草植物首先被采食, 为一些个体较小的非优势种尤其是杂类草功能群的生长提供了较好的微环境, 加速其生长, 使得暖季放牧草地有相对较高的植被密度(图1A)。本研究中放牧草地的一些豆科类植物和杂类草, 例如狼毒(Stellera chamaejasme)、甘肃马先蒿(Pedicularis kansuensis)、黄花棘豆(Oxytropis ochrocephala)等毒杂草也具有很强的繁殖能力, 耐践踏的同时又很少被家畜采食, 可以在暖季放牧中大量地繁殖生长, 因此在暖季放牧草地中具有较高的植物密度, 特别是杂类草密度显著高于冷季放牧草地的(图1B)。暖季放牧虽然一定程度上激发了植物的补偿性生长, 但并不能弥补家畜采食对植物光合能力造成的影响, 地上生物量不能及时再生[25], 这也解释了本研究中暖季放牧草地地上生物量小于冷季放牧草地的结果。在冬季放牧草地中, 放牧家畜采食了大量地上枯落物, 为植物后来的生长提供了充足的光照和空间资源, 有利于植物再生和幼苗的形成, 从而进一步增加了植物的生长量。因此, 冷季放牧草地群落地上生物量显著高于暖季放牧草地的(表2)。
| 表2 不同季节放牧草地各植物功能群生物量(g· m-2)变化 Table 2 Biomass (g· m-2)of different functional groups in seasonal grazing grassland |
放牧季节是影响草地生产力和多样性稳定的重要因素之一, 需要建立合理的放牧制度才能发挥季节性放牧的优势及对草地的调控作用。暖季放牧草地的物种多样性和均匀度指数明显高于冷季放牧草地, 而冷季放牧草地的丰富度指数高于暖季放牧草地。冷季放牧草地的地上生物量以及莎草类和豆科类功能群的生物量比例均高于暖季放牧草地。暖季放牧草地较冷季放牧草地有相对较高的植被密度。合理的放牧强度下, 暖季放牧相比冷季放牧有利于草地群落物种多样性和均匀度的维持, 但冷季放牧草地有利于草地群落地上生物量的积累。对此, 还需要深入探索家畜与草地之间长期的内在关系, 确定合理的放牧制度达到放牧生态系统中动物与植物的协调, 维系草地群落物种多样性和生产力的稳定, 以达到放牧草地生态系统的可持续发展。
The authors have declared that no competing interests exist.
| [1] |
|
| [2] |
|
| [3] |
|
| [4] |
|
| [5] |
|
| [6] |
|
| [7] |
|
| [8] |
|
| [9] |
|
| [10] |
|
| [11] |
|
| [12] |
|
| [13] |
|
| [14] |
|
| [15] |
|
| [16] |
|
| [17] |
|
| [18] |
|
| [19] |
|
| [20] |
|
| [21] |
|
| [22] |
|
| [23] |
|
| [24] |
|
| [25] |
|