


{kind=link}
{kind=link}
{kind=link}
4种豆科牧草种子对绵羊消化道作用的反应
[娜丽克斯·外里 , 王树林, 赵刚, 杨洁晶, 吕艳萍, 鲁为华]
, 王树林, 赵刚, 杨洁晶, 吕艳萍, 鲁为华]
, 王树林, 赵刚, 杨洁晶, 吕艳萍, 鲁为华]
|
|


第一作者:娜丽克斯·外里(1989-),女(哈萨克族),新疆新源人,在读硕士生,主要从事草地资源与生态研究。E-mail:[email protected]
以4种野生豆科牧草苦豆子( Sophora alopecuroide)、线叶野豌豆( Vicia tenuifolia)、光果甘草( Glycyrrhizae glabrae)、罗马苜蓿( Medicago romanica)种子为研究对象,测定各种子在绵羊消化道作用前后的形态、生物学特性变化以及在绵羊消化道内的运移和排放规律。结果表明,4种种子排放过程均符合高斯模型,且苦豆子、线叶野豌豆、光果甘草排放峰值均出现在饲喂后24~36 h,罗马苜蓿种子排放峰值则出现在18~30 h。苦豆子、线叶野豌豆、光果甘草和罗马苜蓿种子总回收率分别为14.513%、4.463%、10.811%、4.608%,在绵羊消化道内的平均滞留时间分别为36.3、29.6、31.2和31.9 h。除苦豆子外,其它3种植物种子在经过消化道后单粒重显著降低( P<0.05),4种种子消化后的形状指数较经过消化道前显著减小。经过绵羊消化道后种皮颜色加深,纹理模糊,出现褶皱及明显侵蚀斑。与经过消化道前比较,4种植物种子经过消化道后的发芽率均有显著提高,其中发芽率提高效果最明显的时间段在消化24~48 h后。消化道作用可有效打破苦豆子、线叶野豌豆、光果甘草、罗马苜蓿种子的硬实,4种植物种子均可通过绵羊消化道传播。
Seeds were collected from four Leguminous plant species ( Sophora alopecuroide, Vicia tenuifolia, Glycyrrhizae glabrae, Medicago romanica) and fed to sheep, then recorded the seeds recovery rates and mean retention time after digestion. Determined the morphological and biological characteristics and germinability of seeds before and after digestion. The results showed that many seeds were excreted from sheep digestive tract, and excretion dynamic all fitted Gaussian model, excretion peak of S. alopecuroide, V. tenuifolia, G. glabrae appeared 24~36 h after digestion, but that of M. romanica appeared 18~30 h after digestion. Total recovery rate from feces of four plants species seeds was 14.513%( S. alopecuroide), 4.463%( V. tenuifolia), 10.811%( G. Glabrae), 4.608% ( M. romanica), respectively. Mean retention time of S. alopecuroide, V. tenuifolia, G. glabrae and M. romanica was 36.3, 29.6, 31.2, 31.9 h, respectively. Seed weight decreased significantly ( P<0.05) after digestion except for S. alopecuroide, seed shape index of all species decreased. The color of seed coat was deepened, seed surface texture was indistinct, and appeared wrinkles and obvious erosion after digested obviously. The germination rates of seed increased after digestion and the highest germination rate appeared in 24~48 h after digestion. Gastrointestinal tract of sheep can break hard seeds and provide a good pathway for seed dispersal.
草食动物对种子的消化道传播是实现种子远距离传播的重要途径之一。种子被草食动物采食营养体吞咽进入消化道, 在游走过程中种子随粪便排出体外, 从而使种子远离母株, 减少了同胞竞争的风险[1]。在动物体内的滞留使得某些具有休眠特性的种子其休眠被打破, 提高了种子的萌发率[2]。同时, 粪便又可为种子萌发和幼苗的生长提供良好的安全地(safe sites)[3]。因此, 草食动物对种子的消化道传播不仅影响种子本身, 而且对于幼苗建成和种群的自然更新具有重要的促进作用, 甚至在更大尺度上改变植物群落的结构与功能[4], 这在过度放牧造成的退化草地上体现的更为明显[5]。
自Janzen[6]提出“ 双F假说” 以来, 草食动物对植物种子消化道传播成为人们广泛关注的焦点。而放牧生态系统中, 家畜和草地植物之间的相互作用更为明显。因此, 近年来, 有关放牧家畜对植物种子消化道传播的影响也备受关注, 目前为止已经涉及到了包括绵羊[7]、牛[8]、牦牛[9]、山羊[10]、驴[11]、马[12]等放牧家畜, 具体研究内容包括种子形态学特征和经消化道作用后的种子回收率、种子活力以及消化道滞留时间之间的关系[13], 种子经消化道作用后萌发幼苗的生长状况以及对植物种群更新过程的影响等方面[3], 并且可进一步基于上述研究内容探讨植物种子传播过程与草食动物之间是否存在协同进化关系[14]。尽管前人已经在种子消化道传播方面做了大量工作, 但基于某一特定类群植物种子在被动物消化后的性状变化以及种子的平均滞留时间、排放规律等方面的研究仍比较缺乏, 而这是开展植物种子传播和动物之间是否存在协同进化关系的基础。在草地生态系统中, 豆科植物是除禾本科外的第二大类群, 并且是草食动物优先选择采食的对象[15, 16, 17, 18]。天然草地上豆科植物多为草本, 且种子与植物叶片、茎秆等可食部分在空间位置上联系紧密, 这为种子被家畜采食并通过消化道传播创造了便利条件。另外, 种子性状是影响消化道传播效率的关键因素[3], 而野生豆科牧草的种子大小、形状在物种之间存在巨大差异, 并存在普遍的休眠现象。因此, 探讨豆科植物种子经消化道作用后的形态学特征、萌发率、种子在消化道内的滞留时间及种子排放规律等, 对于明确豆科植物种子消化道传播的内在机制具有重要意义。
基于上述原因, 本研究以新疆常见的4种豆科植物种子作为研究对象, 采集种子并定量饲喂绵羊, 利用阶段性全收粪法进行种子回收, 并观测种子生物学特性。主要明确以下3个问题:1)不同种子经过消化后的回收率及在消化道内的平均滞留时间; 2)种子消化前后的形态学特征有无变化; 3)种子在消化前后发芽率是否变化? 以进一步丰富草食动物对豆科植物种子消化道传播的研究内容, 并为类似研究提供参考。
选择苦豆子(Sophora alopecuroide)、线叶野豌豆(Vicia tenuifolia)、光果甘草(Glycyrrhizae glabrae)、罗马苜蓿(Medicago romanica)4种豆科植物种子作为研究对象。根据不同牧草种子的成熟时期, 在2012年6月-10月对4种植物种子进行采集。种子采集后于室内风干并置于室外低温条件下(-5~-30 ℃)过冬后置牛皮纸信封常温贮藏。
1.2.1 种子消化道传播实验及在消化道内平均滞留时间 种子饲喂试验从2013年4月开始, 选用体重为(28.0± 2.97) kg的4只健康哈萨克羊, 在具有粪尿分离装置代谢的笼内单笼饲养。饲喂种子前进行为期7 d的预饲, 日粮组成为精料0.2 kg· d-1, 初花期苜蓿(M. sativa)干草1.8 kg· d-1, 自由饮水, 进行试验前两周不饲养任何带有种子的牧草, 预饲期结束后, 保持日粮组成不变, 开始饲喂种子。每只羊每次定量饲喂一种种子2 000粒, 一种植物种子的饲喂及绵羊粪便收集工作完成3 d后, 再饲喂下一种植物种子。种子饲喂后, 在6、12、24、36、48、72 h定时利用阶段性全收粪法收集粪便[19], 所收集粪便在室内常温下静置晾干, 人工搓碎粪便取出消化后的种子, 用自来水冲洗干净后, 计算回收种子数量, 并按公式(1)计算平均滞留时间[20]。
式中:mi指的是动物在摄入种子的ti时刻第i次排便时所释放的种子数, n为排便次数。
1.2.2 种子形态学指标测定 用1/10 000电子天平称量种子的重量, 用体视显微镜测量种子长(L)、宽(W)、高(H)三维指标, 并观察有无附属结构。然后参考Thompson和Hodgson[21]的计算方法根据三维指标计算种子形状指数, 即将种子形状与球体形状对比, 求种子长、宽、高的总体方差, 总体方差的计算方法见公式(2)。方差越小, 种子越接近圆球形; 方差越大, 种子越细长或扁平。在计算方差前要对数据进行转换, 转换方法是先将长定为1, 然后求出宽和高相对于长的比值。
I=[3(
式中:I为种子形状指数, X为各种子的长、宽、高三维指标转换后的数值。
1.2.3 经过消化道前后种子的发芽率 采用纸上法(TP)对从野外采来的种子和经过绵羊消化后的种子进行发芽率检测, 自然状态下种子随机选取100 粒, 消化后种子根据回收数量确定待检测种子数, 除消化后6 h回收种子数量较少不作发芽率检测外, 其它时间段内种子均进行萌发试验, 各植物种子数量在10~100粒, 对于总回收量少于30粒的种子设定两个重复, 其它均为3个重复, 放入双层滤纸培养皿中进行萌发。根据采集地植被生长气候条件设置8 h 15 ℃/16 h 25 ℃变温和光照, 每6、12、24、36、48、72 h进行萌发检测、统计萌发个数, 胚根出现1~2 mm即可认为已经萌发, 直到连续15 d不再有发芽种子出现时为止, 最后以公式(3)计算发芽率。
萌发率=萌发种子数/供试种子数× 100% (3)
| 表1 各时间段不同种子回收率及总回收率(%) Table 1 Recovery rate(%) of seeds from feces after digestion |
苦豆子和罗马苜蓿在饲喂6 h后已有少量种子被回收, 而线叶野豌豆和光果甘草回收率为0(表 1)。饲喂12 h后, 在粪便中均回收到了4种植物种子, 其中以光果甘草和罗马苜蓿种子较多, 回收率分别为1.108%和0.950%, 显著高于其它两种种子(P< 0.05)。饲喂24 h后, 种子回收率明显增加, 其中苦豆子回收率最高, 光果甘草次之, 苦豆子回收率显著高于光果甘草(P< 0.05), 线叶野豌豆和罗马苜蓿回收率较小, 显著低于苦豆子和光果甘草(P< 0.05)。饲喂48 h后, 4种种子回收率明显降低, 至72 h后, 只有极少量种子排出。从4种种子排出数量在各时间段的分布状况看, 罗马苜蓿种子自饲喂6 h后开始排出, 并在后续时间段内排出数量较为均匀。其它3种种子排放时间集中在12~24和24~36 h时间段, 其排放峰值出现在饲喂后的24~36 h。从总回收率看, 苦豆子回收率最高, 达到了14.513%, 显著高于其它3种种子, 罗马苜蓿和线叶野豌豆回收率差异不显著(P> 0.05)。
对4种植物种子在绵羊消化道内的排放动态进行非线性拟合, 为消除不同物种种子回收率之间的量级差异, 在进行拟合之前, 对各物种种子回收率进行Min-max标准化处理。处理后的拟合结果表明(图1), 4物种种子排放过程均符合高斯模型, 其中苦豆子(S)、线叶野豌豆(V)、光果甘草(G)种子排放高峰均在24~36 h时间段, 而罗马苜蓿排放高峰在18~30 h时间段内。
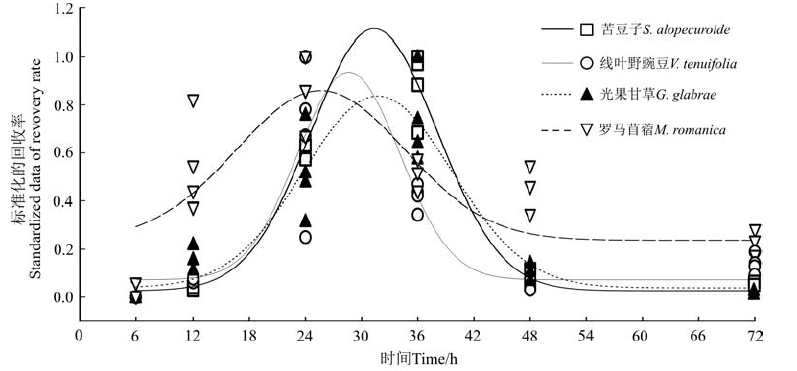 | 图1 4种种子回收率动态Fig.1 Dynamic curves recovery rate and mean retention time of four species seeds in sheep’ s gut |
进一步对各种子在消化道内的平均滞留时间进行计算(图2), 苦豆子的平均滞留时间为36.3 h, 线叶野豌豆为29.6 h, 光果甘草为31.6 h, 罗马苜蓿为31.9 h。其中苦豆子平均滞留时间最长, 显著高于线叶野豌豆(P< 0.05), 线叶野豌豆、光果甘草和罗马苜蓿三者之间差异不显著(P> 0.05)。
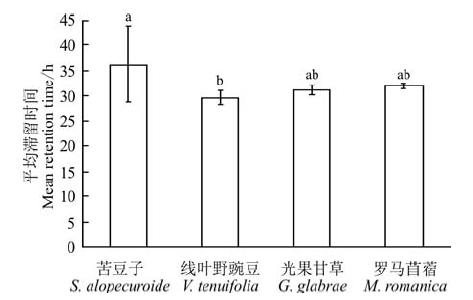 | 图2 4种种子消化道内平均滞留时间Fig.2 Mean retention time of four species seeds in sheep’ s gut |
将经过消化道前、后各时间段的种子在体视显微镜下观察并拍照, 以观察经过消化道前、后的种子的形态学变化。结果表明(图3), 4种植物种子在经过消化道前种皮均很光滑, 表面纹理清晰。而经过消化道后种皮颜色明显加深, 消化36 h后各种子表面出现明显的侵蚀斑。苦豆子种皮在36 h后出现明显褶皱, 并有种子出现种皮开裂和脱落现象。光果甘草在消化48 h后种子表面出现侵蚀斑, 种皮颜色在72 h后明显加深。线叶野豌豆种子在消化6 h后种皮表面纹理可见, 但在12 h后纹理模糊不清, 之后种皮颜色明显加深, 至48 h后种皮表面出现明显侵蚀斑。罗马苜蓿种子在消化24 h后种皮颜色开始加深, 但种子表面侵蚀斑不明显, 种皮相对完整。
 | 图3 种子消化前后形态学特征Fig.3 Morphological difference of seeds before and after digestion |
进一步对4种植物种子经过消化道前后的重量、长、宽、高进行测量, 并计算经过消化道前、后的形状指数, 结果(表2)表明, 除苦豆子经过消化道后单粒重无显著变化外(P> 0.05), 其它3种种子经过消化道后种子单粒重均显著低于消化前(P< 0.05); 4种植物种子经过消化道后种子形状指数均有显著减小的趋势。
| 表2 种子消化前和消化后(36和48 h)形态学指标 Table 2 Morphological feature of seeds before and after digestion (36 and 48 h) |
供试4种种子在经过消化道前其萌发率均较低, 其中线叶野豌豆、光果甘草、罗马苜蓿仅分别为3.33%、5.36%、7.66%, 萌发率较高的苦豆子也仅为12.66%(表3)。经过绵羊消化道后, 种子萌发率均有所提高, 其中苦豆子在消化12 h后, 萌发率提高到43.81%, 并且随着消化时间的延长, 萌发率显著增加, 至48 h后, 萌发率已提高到82.76%, 极显著高于经过消化道前(F=7.981, P=0.001 6)。光果甘草在消化12 h后, 萌发率仅有小幅增加, 但在消化24 h后提高, 至36 h萌发率达到最高的74.26%, 极显著高于经过消化道前(F=75.118, P=0.000 1), 之后则有所下降。罗马苜蓿在经过消化道后萌发率变化与光果甘草相似, 消化36 h后其萌发率达到最大值, 但至48 h后则下降为27.64%, 但总体上消化24~72 h后的萌发率均极显著高于消化前(F=113.414, P=0.000 1)。线叶野豌豆经过消化道后, 萌发率在12和24 h仅有小幅提高, 至36 h后则极显著提高到44.51%(F=106.112, P=0.000 1), 但提高幅度显著低于其它3种牧草(F=73.06, P=0.001)。
| 表3 种子消化前和消化各时间段萌发率(%) Table 3 Germination percentage(%) of seeds in every time after digestion |
以物种和消化道停留时间两个因素进行方差分析(表4), 不同物种对发芽率影响极显著(P< 0.01), 消化不同时间对种子发芽率影响也极显著(P< 0.01), 物种和不同消化时间两个因素间互作效应也极显著(P< 0.01)。
| 表4 物种和消化时间对种子发芽率影响的双因素方差分析 Table 4 Results of two-way ANOVA of effect of species and digestion time on seeds germination percentge |
前人的研究结果表明, 缺乏助力传播结构特征的种子由于传播距离非常有限, 往往需要通过动物携带来实现远距离传播, 而草食动物会将大量种子与植物营养体一起采食并吞咽至消化道内, 为种子的远距离传播提供了另一个有效途径[6]。本研究中除了线叶野豌豆种子以外的其它3种豆科植物种子由于缺乏特化传播结构使传播距离受到限制[22], 而动物消化道传播为长距离传播提供了便利条件, 但要成功实现消化道传播, 必须要满足两个条件:1)有足够数量且具生活力的种子能够成功随粪便排出动物体; 2)种子在消化道内要有充足的滞留时间, 以便于通过畜体携带游走而产生大尺度位移[22]。豆科植物Retama sphaerocarpa和Cytisus scoparius的种子在饲喂绵羊后其排放时间段集中在20~36 h, 回收率在12%~16%[7]。白花三叶草(Trifolium repens)种子在绵羊体内的平均滞留时间为51 h左右, 这段时间足够种子被远距离传播[23]。本研究也得到了类似的结果, 4种植物种子在饲喂绵羊后, 自12 h时间段开始, 就有大量种子排出, 并在18~36 h时间段排出数量达到高峰, 种子总体回收率在5%~15%。同时, 种子在绵羊体内的平均滞留时间在29~36 h。而Cory[24]的研究结果表明, 自由放牧绵羊每天的游走距离在6.1 km, 充分说明这4种植物种子被放牧绵羊采食后, 可伴随畜体移动游走有充足的时间实现远距离传播, 从而对物种种群更新和拓展新生境提供了有效途径[25]。
种子在草食动物消化道内的回收率和运移过程与种子的形态学特征密切相关, 主流观点认为, 圆形种子通过消化道的速度要快并且回收后的萌发率要高[8]。因此, 有学者提出了一个种子形状指数的概念(shape index), 认为种子形状指数越小(即种子越接近圆形), 其通过消化道的速度越快[26]。这一观点在豆科牧草和禾本科牧草种子经过消化道传播的相关试验中已经得到了很好的验证[27, 28]。本研究中的4种豆科植物种子的形状指数均小于0.1, 为典型的圆形种子[21], 因此, 一旦从动物的咀嚼过程中“ 逃逸” 进入消化道后, 可相对安全快速地通过消化道并被排出体外而实现传播。但是, 就同一形状指数水平的种子来说, 种子大小又是影响种子在消化道内滞留时间的关键因素。已有研究结果表明, 小种子除了在咀嚼过程中能够成功“ 逃逸” 之外, 其在动物消化道的通过速度也很快[29], 并且被反刍破坏的几率小[30]。但经过长期的试验研究发现, 种子大小和消化道传播之间并非简单绝对的线性相关关系。种子在消化道内的排出速率在其大小规格存在一个临界值, 低于这一临界值的种子经过家畜消化道的速度可能很快, 即通过速率并非完全随着种子体积缩小而单调增加, 而是基本保持在稳定的水平, 并着重指出低于这一临界值的小种子可以很容易通过瘤网口, 因此其存活可能性就高。反之, 如果高于这一临界值的话, 则速度可能会变慢[14]。本研究中的罗马苜蓿虽然是典型的圆形小种子, 但其在消化道内的平均滞留时间为31.96 h, 并未显著低于其它3种种子, 说明该小型种子大小规格并未达到快速排出的临界值。
动物消化过程会对种子重量、大小产生显著影响。高寒草甸中的多种植物种子在被藏羊和牦牛消化后, 种子重量显著减小, 并且会影响到幼苗的生长[9]。本研究结果也得到了类似的结论, 经过消化后有3种植物种子重量显著降低, 且所有种子的形状指数明显减小。到目前为止, 关于消化作用对种皮颜色和纹理影响的报道仅在鸟类的种子消化道传播方面[31], 而在草食动物的种子消化道传播方面未见报道, 而本研究发现, 随着消化时间增加, 种皮颜色加深、纹理模糊, 甚至有种皮开裂, 因此, 对由于特化种皮结构而引起物理性休眠的豆科植物种子, 种皮表面结构特征在消化前后的变化也是今后需要关注的问题。
豆科植物种子普遍存在物理休眠现象, 种子在动物消化道内的运移过程直接影响到种子的萌发。众多研究表明, 具有休眠特征的种子在经过动物消化道作用后排出体外, 其休眠特征均被不同程度地打破[32], 除了种子经过啃咬和咀嚼可导致部分种皮被破坏, 引起种子透水、透气性增加外[33], 主要原因是种子在动物消化道内瘤胃消化液、微生物和众多酶类的综合作用下, 种皮软化, 透气透水性增强, 从而破除了种子的休眠[9]。本研究的4种豆科种子消化前的萌发率均较低, 但经过消化后其发芽率显著提高, 结合种皮表面形态特征变化, 充分说明由种皮造成的物理性休眠被消化过程所打破, 从而使种子的发芽率显著增加。但是, 从本研究中也发现, 4种植物种子在被消化72 h后, 其发芽率不再增加, 反而有减小的趋势, 因此。种子在消化时间和休眠解除两方面又存在明显的“ 权衡” , 如果消化时间过长, 则会对种子本身产生破坏作用, 有可能使种子活力丧失。
4种植物种子在饲喂绵羊12~72 h各时间段均有种子被排出, 其总体回收率在5%~15%, 其中以苦豆子和光果甘草回收率较高。4种种子排放过程均符合高斯模型, 其中苦豆子、线叶野豌豆、光果甘草种子排放高峰均在24~36 h时间段, 罗马苜蓿种子排放高峰在18~30 h时间段。各种子在绵羊体内的平均滞留时间在29~36 h, 其中苦豆子种子在消化道内的平均滞留时间最长, 线叶野豌豆种子的平均滞留时间最短, 光果甘草和罗马苜蓿居于二者之间。
种子经过消化道后, 除苦豆子外, 其它3种种子重量显著减小, 所有种子形状指数均明显减小。种皮表面有明显侵蚀斑、裂纹, 甚至种皮脱落, 纹理模糊。经过消化道后, 发芽率提高效果最明显的时间段在消化24~48 h后, 不同程度地解除了种子的休眠, 并且各种子由于在畜体内有较长的滞留时间, 从而能够通过畜体位移实现有效的远距离传播, 为4种植物种群更新过程中减少同胞竞争及增加拓殖生境的多样性提供了良好条件。
The authors have declared that no competing interests exist.
| [1] |
|
| [2] |
|
| [3] |
|
| [4] |
|
| [5] |
|
| [6] |
|
| [7] |
|
| [8] |
|
| [9] |
|
| [10] |
|
| [11] |
|
| [12] |
|
| [13] |
|
| [14] |
|
| [15] |
|
| [16] |
|
| [17] |
|
| [18] |
|
| [19] |
|
| [20] |
|
| [21] |
|
| [22] |
|
| [23] |
|
| [24] |
|
| [25] |
|
| [26] |
|
| [27] |
|
| [28] |
|
| [29] |
|
| [30] |
|
| [31] |
|
| [32] |
|
| [33] |
|