

{kind=link}
{kind=link}
{kind=link}
兴隆山自然保护区华北珍珠梅物候期对水热条件的响应
[潘世成1  , 祁军
, 祁军1 , 张学炎1 , 陈蕾1 , 王静2 , 孟秀祥2 ]
, 祁军]
|
|


第一作者:潘世成(1965-),男,甘肃临洮人,高级工程师,本科,主要从事自然保护区科研工作。E-mail: [email protected]
华北珍珠梅( Sorbaria kirilowii)为蔷薇科珍珠梅属植物,具有抗病虫害等生态价值,是兴隆山自然保护区内主要的灌木种。本研究通过对甘肃省兴隆山国家级自然保护区内华北珍珠梅2004-2007年及2012-2015年9种物候特征的观察,分析了华北珍珠梅各物候期的变化趋势以及各物候期与月平均气温和月降水量的相关关系。结果表明,华北珍珠梅的春季物候期多数呈现推迟趋势,而秋季物候期主要呈现提前趋势,整个生长季的时间每年缩短达15.49 d。华北珍珠梅物候期的变化趋势与月平均气温和月降水量具有密切的相关关系,其中开花期和果实膨大期与4月份的平均气温间呈显著的负相关关系( P<0.05),发芽期与2月份降水量,抽枝期与5月份降水量以及果实膨大期与7月份降水量均呈显著负相关关系( P<0.05),而果实成熟期与10月份降水量呈显著的正相关关系( P<0.05)。因此,受水热条件影响的华北珍珠梅生长季的缩短不仅会降低其生长繁殖,也会减少灌木层的郁闭度,不利于生活在灌木层的动物,如濒危物种马麝( Moschus sifanicus)等的栖息。
Sorbaria kirilowii belongs to Rosaceae, with obvious ecological value such as resistant to diseases and insects, which is also a main shrub species in Xinglongshan Natural Reserve. Through two periods, 2004-2007 and 2012-2015, nine types phenologic phase of S. kirilowii in Xinglongshan National Natural Reserve were observed to analyze the change tendency of phenologic phase of S. kirilowii and explore the relationships of phenologic phase to monthly mean temperature and precipitation. The results indicated that most Spring phenologic phases for S. kirilowii showed a trend of delay but an advanced trend of Autumn phenologic phases, which caused the decrease of growth period, as much as 15.49 d·a-1. The trend of phenologic phases could be resulted from the phonologic phases of S. kirilowii correlated significantly with monthly mean temperature and precipitation. The blooming and fruit expansion periods are significantly negative correlated with mean temperature of April. There are significantly negative correlations between budding period and the precipitation of February, branching and the precipitation of May, fruit expansion period and the precipitation of July, while there is significantly positive correlation between fruit ripe period and the precipitation of October. Growth decrease caused by regional climate change influence not only propagation, but brush canopy density, which against animals living, such as Moschus sifanicus.
物候是指生物长期适应温度条件的周期性变化, 形成与此相适应的生长发育节律, 主要包括动植物的生长、发育、活动规律与非生物的变化对物候的反应[1]。在全球气候变化、生态问题日益突出的背景下, 植物物候对气候变化的响应及其模拟已成为一个研究热点[2, 3]。近年来的研究表明, 在众多影响植物物候的环境因子中, 温度起主要作用[4], 随着研究的进一步深入, 目前植物物候主要表现为随温度升高, 春季物候期提前, 秋季物候期推迟, 生长期相对延长[5]。除了温度外, 降水量是影响植被生长发育最重要的要素之一。降水对植物物候的变化也会产生影响。例如, 海南岛尖峰岭12种热带常绿阔叶乔木在展叶和开花期受气温和降水综合影响最明显的月份, 若其月平均气温升高0.1 ℃、月降水量增加10 mm可使展叶始期和开花始期提前或推迟1~3 d[6]。
兴隆山地处青藏高原、黄土高原、蒙新高原交汇地带, 区内海拔跨度大、生态系统类型齐全, 濒危动植物及特有植物物种极多, 是我国的生物多样性中心及重要基因库[7]。其中, 华北珍珠梅(Sorbaria kirilowii)为蔷薇科珍珠梅属植物, 具有花期长、耐寒、耐半阴、繁殖容易等优良特性。华北珍珠梅作为兴隆山自然保护区内主要的灌层物种, 具有较高的郁闭度, 为生活在灌层的动物如濒危物种马麝(Moschus sifanicus)提供了理想的隐蔽场所。此外, 华北珍珠梅对烟尘等有害气体有不同程度的吸收和抗性, 并能产生挥发性的植物杀菌素, 杀菌效果显著且稳定[8]。这一杀菌作用对防治高山灌丛和草甸中的有害微生物的污染具有重要的意义。目前, 对华北珍珠梅的物候已有一些研究结果[9], 但对该地区的相关研究较少。因此, 针对兴隆山保护区内5个分布点华北珍珠梅开展物候期与气候变化的研究, 进而明确该地区华北珍珠梅物候变化趋势及对气候变化的响应, 以期对预测保护区内华北珍珠梅的物候期并开展适当的保护、保育工作, 以及对了解保护区内灌木层郁闭度的变化动态及其动物栖息环境的影响提供一定的参考作用。
兴隆山自然保护区位于103° 50'-104° 10' E, 35° 38'-35° 58' N, 属祁连山的东延余脉, 总土地面积为29 583.6 hm2, 年总径流量5 349 m3, 全区森林覆盖率82.5%, 活立木总蓄积量60.6万m3, 海拔1 800-3 670 m, 属大陆性季风气候, 总的气候特征表现为温带半湿润气候。保护区日照适中, 降水充足, 气温偏低。兴隆山自然保护区是森林生态系统类型的自然保护区, 主要保护对象为野生动物马麝(国家Ⅰ 级)和以青杄(Picea wilsonii)、云杉(Picea asperata)为主的水源涵养林及其生态系统。其中, 大果圆柏(Sabina tibetica)、紫果云杉(Picea purpurea)、巴山冷杉(Abies fargesii)和榆中贝母(Fritillaria yuzhongensis)等是具有重要保护价值的国内特有种。马麝作为保护区主要受保护的哺乳纲动物, 华北珍珠梅等灌木物种为其提供理想的栖息场所。
选取长势良好、连续3年没有病虫害的华北珍珠梅, 对其发芽期、芽膨大期、展叶期、开花期、结果期、果实膨大期、抽枝期、果实成熟期和落叶期9种物候现象进行观察纪录。观测点分布在保护区各管理站(表1)。
| 表1 观测点分布表 Table 1 Observation point distribution |
气象资料来源于中国气象科学数据共享服务网(http://data.cma.cn), 包括2004-2015年日最高温、日最低温、日平均温度以及日降水量。
观测方法参照《中国物候观测方法》[10], 自2004-2007年及2012-2015年, 观察记录华北珍珠梅的9种物候现象, 即发芽、芽膨大、展叶、开花、结果、果实膨大、抽枝、果实成熟、落叶。自每年3月中旬起, 每天进行观察记录, 将观测对象50%出现变化时, 规定为该物候现象的物候期。
数据处理中, 采用儒略日(Julian days)换算方法将物候期观测数据转化为距当年1月1日的天数, 从而得到华北珍珠梅各个物候现象的日序。
采用SPSS软件对华北珍珠梅不同年份各物候期进行回归分析以探讨兴隆山保护区每年9个观测的物候期的变化趋势。使用Spearman相关分析法分析9种物候期、月平均气温和月降水量的相关关系, 以探讨物候期对水热条件变化的响应。
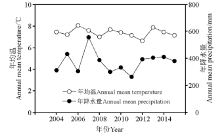
从2004-2015年, 兴隆山地区年均气温为7.37 ℃, 年均最高温为15.07 ℃, 年均最低温为1.19 ℃, 年均降水量为372.77 mm。近12年来, 年均气温呈下降趋势, 而年降水量呈上升趋势(图1)。
 | 图1 兴隆山保护区2004-2015年年均气温和降水变化趋势Fig.1 The trends of annual mean air temperature and precipitation from 2004 to 2015 in Xinglongshan Natural Reserve |
华北珍珠梅物候期的发生顺序相对较为稳定。其中发芽、芽膨大和展叶期主要发生在3和4月, 5-7月主要为开花和结果期, 8-10月主要为果实成熟和落叶期, 而抽枝期在不同年份间的变异较大(图2)。
 | 图2 兴隆山保护区华北珍珠梅物候时间序列及物候变异分位数分布图 注:P1, 发芽期; P2, 芽膨大期; P3, 展叶期; P4, 开花期; P5, 结果期; P6, 果实膨大期; P7, 抽枝期; P8, 果实成熟期; P9, 落叶期。Fig.2 Phenological time series of Sorbaria kirilowii and variability of data in quartiles in Xinglongshan Natural Reserve Note: P1, Budding period; P2, Bud expansion period; P3, Leaf unfolding period; P4, Blooming period; P5, Fruiting period; P6, Fruit extension period; P7, Branching period; P8, Fruiting ripe period; P9, Leaf-fall period. |
华北珍珠梅不同年份各物候期回归分析结果表明(图3):发芽期、芽膨大期、展叶期和抽枝期均有推迟的趋势, 其中, 芽膨大期每年约推迟3.97 d; 而开花期、结果期、果实膨大期、果实成熟期和落叶期均表现出提前的趋势, 其中, 果实成熟期每年约提前3.83 d。总体来看, 华北珍珠梅除开花期外, 其它观测到的所有春季物候期均呈现推迟的趋势, 而所有秋季物候期均出现提前的趋势。华北珍珠梅春季物候期平均每年推迟4.26 d, 秋季物候期平均每年提前11.23 d, 即生长季节每年平均缩短15.49 d。
 | 图3 2004-2007年和2012-2015年华北珍珠梅各物候期变化趋势Fig.3 Trend changes of phenophases of Sorbaria kirilowii from 2004 to 2007 and 2012 to 2015 |
在植物生长发育的各物候期中, 物候的开始日期与其前期气温之间具有密切的关系[8], 通过Spearman相关分析得出华北珍珠梅各物候期与各月均温间的相关系数(表2)。华北珍珠梅属于晚春到初夏开花的灌木, 其开花期和果实膨大期与4月份的平均气温间呈显著的负相关关系(P< 0.05), 即随着温度的升高, 开花与果实膨大将提前。这说明, 4月份温度的高低是影响华北珍珠梅开花早晚的关键。虽然发芽期与1月、2月和3月平均气温间并未达到显著相关水平(P> 0.05), 但呈负相关关系, 也表明温度升高, 发芽也将随之提前。
| 表2 华北珍珠梅各物候期与月平均温度的相关性分析 Table 2 Correlation between phenological phases of Sorbaria kirilowii and monthly mean temperatures |
华北珍珠梅发芽期与2月份降水量呈极显著的负相关关系(P< 0.01)(表3)。这说明发芽前降水的增加有助于促进华北珍珠梅提前开花。华北珍珠梅果实膨大期主要发生在7月份, 其与当月降水量呈显著的负相关关系(P< 0.05), 说明当月降水量对果实膨大起着关键作用。华北珍珠梅果实成熟期与10月份降水量呈显著的正相关关系(P< 0.05), 这说明10月降水量的增加将抑制果实的成熟。
| 表3 华北珍珠梅各物候期与月降水量的相关性分析 Table 3 Correlation between phenological phases of Sorbaria kirilowii and monthly precipitation |
物候期变化主要受温度的影响[11]。近60年来, 我国地表平均气温每10年上升0.23 ℃, 气候变暖的幅度明显高于全球。研究表明, 与1986-2005年相比, 2016-2035年全球地表平均气温预计将升高0.3~0.7 ℃; 至21世纪末将升高0.3~4.8 ℃, 我国将平均升高1.3~5.0 ℃[12]。国内外的许多研究都报道了春季物候提前、秋季物候延迟的现象[5]。例如, 欧洲春季物候如树木展叶时间提早6 d[13]。北美地区1959-1993年丁香(Syzygium aromaticum)春季展叶时间提前5.4 d, 发芽提前4.2 d[14]。在美国马萨诸塞州伍斯特郡地区过去50年的气候数据显示春季温度平均每10年上升0.24 ℃, 平均花期提前2.9 d[15]。中国温带地区植物在1982-1999年间返青日期平均每年提前0.79 d[16]。哈尔滨市在1963-2008年气温呈显著上升趋势, 华北珍珠梅的芽开放期、展叶始期和开花始期都呈提前趋势, 而脱落末期则推迟[8]。本研究中, 兴隆山保护区在2004-2015年的气温呈下降趋势。华北珍珠梅物候的研究结果显示, 春季物候期呈推迟趋势, 秋季物候期则主要呈现提前趋势。这一物候变化趋势与温度和降水的变化密切相关。虽然这一趋势与目前多数研究得到的变化趋势相反, 但也从反面验证了温度上升导致春季物候期提前的研究结果。植物物候期受到遗传和环境因素的复杂影响, 其中水热条件可能直接控制着物候的开始时间[17]。华北珍珠梅物候期的变异是不同个体中表现出的遗传变异, 还是个体对环境异质性的可塑性响应还需进一步的研究。
温度和降水对植物生长发育具有重要的作用。其中对植物生长具有直接作用的热量指标和降水量分别为有效温度和供给植物根系的土壤水。张学霞等[18]提出, 在进行年际变化分析不同月份的温度和降水与物候的关系时, 完全可以只讨论实际气温、实际降水指标与植物物候之间的关系。本研究中, 兴隆山地区华北珍珠梅的春季物候推迟的现象与温度的变化具有一定的负相关关系。但由于本研究物候观测数据量相对较少, 未能发现物候期与温度变化的显著相关性。植物物候对温度变化的响应是一个复杂的生理过程。随着气温的增加, 物候期提前的幅度却逐渐减少, 当温度增加的幅度减少, 物候推迟的程度却增加[19]。例如, 对欧洲1 245个观测点、7种关键树种的长期观测发现:在1980-2013年, 温度每上升1 ℃, 春季展叶的提前天数减少, 平均提前天数从1980-1994年的(4.0± 1.8)d· ℃-1减少到1999-2013的(2.3± 1.6)d· ℃-1[19]。华北珍珠梅对温度变化的响应还需要更多观测数据的积累, 有待于进一步的分析探讨。
除了温度外, 降水对植物物候的变化也会产生影响。例如, 对葡萄牙地区广布的入侵种相思草(Acacia longifolia)的研究发现, 虽然温度对其物候现象有主要作用, 但降水对发芽、开花的物候现象有更为显著的影响[20]。尤其是在夏秋交替季节的热带地区, 由于温度和光周期变化不明显, 植物物候受水分的影响很显著[21]。本研究中, 兴隆山地区的降水量的季节分布极不均匀, 降水量主要集中在夏季, 春、秋两季降水量较少。降水对华北珍珠梅的发芽、果实膨大、果实成熟具有显著不同的影响。其原因可能是, 2月份降水量的增加会提高土壤湿度, 加速植物的生长, 表现在春季发芽提前。果实膨大对水分高的需求也导致该物候期与当月实际降水量间的显著相关性。10月份降水量的增加使得华北珍珠梅果实成熟推迟。降水与温度对华北珍珠梅物候的综合作用需要进一步深入的研究。
兴隆山地区华北珍珠梅的生长期减短, 不利于其生长繁殖。另一方面, 植被物候期的变化会对保护区其它物种的生长、存活也产生重要的影响。以马麝为例, 郁闭度是影响兴隆山马麝秋季生境选择的重要因素[22], 冬季马麝更喜欢选择针阔混交林和灌丛[23], 而华北珍珠梅的生长季缩短, 会导致保护区内灌木层的郁闭度在春初秋末时期降低, 影响以濒危马麝为代表的主要以灌木层为隐蔽场所的动物的栖息。由于植物物候本身受多种因素的影响, 且较短时间的观测以及未对其它气候因素进行综合分析可能会对物候的准确性判断产生影响, 因此长时间段的物候监测是未来需进一步研究的主要任务。
The authors have declared that no competing interests exist.
| [1] |
|
| [2] |
|
| [3] |
|
| [4] |
|
| [5] |
|
| [6] |
|
| [7] |
|
| [8] |
|
| [9] |
|
| [10] |
|
| [11] |
|
| [12] |
|
| [13] |
|
| [14] |
|
| [15] |
|
| [16] |
|
| [17] |
|
| [18] |
|
| [19] |
|
| [20] |
|
| [21] |
|
| [22] |
|
| [23] |
|