



{kind=link}
{kind=link}
{kind=link}
{kind=link}
海口城市草坪绿地土壤CO2通量时间变化及其环境解释——以海南师范大学南校区为例
[徐文娴1  , 卓志清
, 卓志清2 , 赵从举1 , 朱敏捷1 , 韩奇1 , 吴小卫1 ]
, 卓志清, 朱敏捷|
|


第一作者:徐文娴(1987-),女,甘肃西和人,硕士,主要从事土壤资源生态利用研究。E-mail:[email protected]
利用LI-8100A土壤碳通量测量系统对热带城市草地土壤CO2通量进行定点系统监测,阐明土壤CO2通量时间变化特征及其对环境因子响应。结果表明,1)草坪绿地土壤CO2通量日变化呈单峰曲线,最高值一般出现在09:00-16:00,不同月份、不同天气条件有所差异;土壤CO2通量的日平均值6月份最大,为4.27 μmol·(m2·s)-1,12月份最小,为0.86 μmol·(m2·s)-1,年内变化趋势总体呈单峰曲线;2)草坪绿地土壤CO2通量变化与气温、土壤温度呈显著的正相关关系( P<0.05),而与土壤体积含水量之间的关系较为复杂,相关性不显著( P>0.05)。3)较小强度降水对土壤CO2通量变化影响较小,而强降雨对土壤CO2通量变化产生明显的挤出作用。一次大的降水过程初期,随着降水量增加,土壤呼吸速率持续增大;而当土壤水分含量较高时,降雨则对土壤呼吸产生抑制作用。
To explore the change characteristics of soil CO2 flux and its response to air temperatures, soil temperatures, soil moistures and other environmental factors, the soil respiration rates of tropical urban lawn ecosystems were measured by using the LI-8100A Soil CO2 Flux System, and meanwhile air temperatures, soil temperatures at 5 cm and soil moisture 0-10 cm in depth were monitored from November of 2013 to October of 2014. The results showed that: 1) Daily variation of soil respiration was single peak curve. The peaks usually occurred at 09:00-16:00, and the time when peak values generally appeared were different in different months and weather conditions. Yearly variation of soil respiration was also a single peak curve. The mean daily soil respiration rate was 0.86 to 4.27 μmol·(m2·s)-1, with the maximum in June, and the minimum in December. 2) There was a significant positive correlation between soil respiration rate and air temperature and soil temperature. However, the relationship between the CO2 flux and soil volumetric water content was more complex, and the correlation was not significant. 3) Rainfalls had complicated effects on soil respiration. A light rain didn’t significantly change soil respiration, but a heavy rainfall could have an obvious extrusion effect on soil CO2 flux. At the early stage of a heavy rainfall process, soil respiration rate increased with the increase of rainfall, however, continuous rainfall could increase the soil moisture, and rainfall became an important factor limiting soil respiration.
土壤呼吸是土壤有机碳向大气输出的主要途径, 是地表土层与近地层大气之间碳交换的数量表征[1]; 土地利用变化所引起的土壤CO2净排放量大约相当于化石燃料所释放的CO2排放量的1/4, 成为大气CO2浓度增加的第二大人为碳源[2]。土壤CO2释放研究对估算未来大气CO2浓度及全球气候变化具有举足轻重的意义[3]。目前, 土壤CO2通量研究多集中在森林、草地和农田等典型生态系统[4, 5, 6, 7], 而城市生态系统土壤CO2通量研究较少, 热带地区城市绿地土壤呼吸作用未见报道[8, 9, 10]。城市草坪绿地因受到强烈的人类活动影响, 绿地的植被特征和土壤性质与自然草地系统差异较大, 可能导致土壤呼吸过程也存在较大差异[11]。随着城市化进程的加快, 城市用地面积快速扩张, 城市景观绿地土壤CO2通量在全球碳循环中的作用也越来越重要; 而不同区域因气候、土壤以及土壤微生物的差异, 土壤CO2通量呈现出不同规律与特征[12, 13, 14]。
基于此, 从2013年11月起, 利用LI-8100A土壤CO2通量测量系统对热带城市海口的草坪绿地土壤CO2通量进行为期1年的定点原位系统观测, 旨在揭示热带城市草地土壤CO2通量的日年变化特征, 探讨环境因子对土壤CO2通量的影响, 弥补我国热带土壤CO2通量研究资料的缺乏和不足, 为城市生态系统碳收支计算提供基础数据, 为低碳城市建设提供决策参考。
海南师范大学南校区地处低纬度热带北缘, 为热带海洋性季风气候。全年分旱、雨两季, 5月至10月为雨季, 11月至次年4月为旱季; 雨季高温多雨, 旱季温暖少雨; 年均日照超过2 000 h, 年均温23.8 ℃, 年均降水量1 664 mm, 年平均蒸发量1 834 mm; 盛行东南风和东北风, 年均风速3.4 m· s-1。地势平缓, 属滨海平原区, 地表为第四纪基性火山岩和第四系松散沉积物, 土壤为沙质壤土。校区绿地面积占45%以上, 主要包括乔灌结合的行道树绿地和乔灌草结合的花园式绿地。栽种的乔木树种主要有棕榈科、豆科、桑科等, 以棕榈科为主; 灌木与草本植物种类繁多, 主要有芸香科、蔷薇科、紫茉莉科、唇形科、锦葵科、禾本科、马鞭草科、三白草科等[15]。研究样地(19° 59'52″-19° 59'58″ N, 110° 20'18″-110° 20'21″ E)地处海南师范大学南校区, 为花园式城市绿地, 是乔、灌、草相结合的人工植物群落; 乔木层主要有王棕(Roystonearegia)和椰子(Cocos nucifera), 以孤植为主, 覆盖度约20%; 灌木层主要有软叶刺葵(Phoenix roebelenii)、扶桑(Hibiscus rosa-sinensis)、假连翘(Duranta repens), 以丛植为主, 覆盖度约15%; 地被层主要是地毯草(Axonopus compressus)、竹节草(Chrysopogon aciculatus), 绿地植被覆盖度接近100%。布置3个监测样点, 样点间生境相似, 土壤性状比较均一, 管理措施一致。草坪绿地为自然生长, 不翻耕, 不施肥, 不喷药, 仅在旱季进行喷灌式人工浇水, 全年浇水次数视干旱情况而定, 一般为10~20次; 每年采用便携式割草机修剪4~5次, 留茬高度约5 cm; 每次修剪之后, 将杂草、落叶一起运走。样地内表层土壤(0-20 cm)基本性状见表1。
| 表1 样地0-20 cm土壤基本概况 Table 1 The basic soil characteristics of the sampling sites (0-20 cm) |
利用开路式土壤碳通量测量系统 (LI-8100A, LI-COR Inc., Lincoln, NE, USA) 测定土壤呼吸速率。在南校区花园式绿地设置3个监测点, 每个测点安置1个土壤环(内径20 cm, 高12 cm的PVC圆环)。安装土壤环时, 要尽量减少对土壤表层扰动, 并保持基座高出土壤表面3 cm; 为减少对土壤的干扰, 观测期土壤环留在原地。每次实验开始前一天, 将基座内出露地表绿色植物齐地剪掉, 并尽可能不扰动地表凋落物, 不破坏土壤结构, 以减少人为扰动对测量结果的影响。2013年11月-2014年10月每月选取晴好天气2~3 d定点连续监测土壤CO2通量变化, 仪器的测量时间设置为2 min, 重复测量3次, 每隔0.5 h测定一次土壤CO2通量; 同时, 利用仪器配套的土壤温度、土壤水分传感器(TDR)同步测量5 cm深度地温和0-10 cm深度土壤体积含水量, 3个监测点数据取平均值进行统计分析, 气温数据由学校气象站采集。另外, 增加一次雨天(9月7日)土壤CO2通量监测, 从07:00持续观测到19:00, 监测项目同上。
土壤呼吸的温度敏感系数Q10的计算公式为[16]:Q10=e10b。b为温度敏感系数, 是土壤呼吸速率与土壤温度指数模型(Rs=aebT)中的拟合常数, Rs为土壤呼吸速率, a为拟合常数, T为土壤温度。土壤微生物适宜的温度范围一般为16~30 ℃, 结合土壤温度数据, 分别计算旱季(日最高土壤温度< 30 ℃)、雨季(日最高土壤温度> 30 ℃)以及全年Q10。
利用LI-COR提供的数据处理软件和Micrsoft Excel 2007、SPSS 21.0等软件对采集的数据进行分析处理。
2.1.1 草坪绿地土壤CO2通量日变化特征 土壤CO2通量日变化基本呈单峰曲线, 但也存在不规则的波动。选取2013年11月-2014年10月观测期间的1月5日-1月7日、9月26日-9月28日连续3 d的观测值为旱季、雨季土壤CO2通量日变化的代表时段。观测之前天气晴好, 至少连续5 d没有降雨。旱季(1月5日-1月7日)与雨季(9月26日-9月28日)晴天的土壤CO2通量日变化趋势呈现出白天高于夜晚的单峰曲线(图1)。1月5日-1月7日土壤CO2通量日峰值出现在10:00-16:00, 谷值出现在00:30-04:00; 最低值出现在1月6日02:30前后, 为1.08 μ mol· (m2· s)-1, 而最高值出现在1月6日的10:00, 为1.62 μ mol· (m2· s)-1。9月26日-9月28日土壤CO2通量日峰值出现在09:00-11:30, 谷值出现在21:00-次日00:00; 最低值出现在9月29日00:00前后, 为2.12 μ mol· (m2· s)-1, 而最高值出现在9月26日09:00, 为3.36 μ mol· (m2· s)-1。可见, 旱季土壤CO2通量到达峰值、谷值的时间较雨季有一定的滞后性, 且数值也较低。
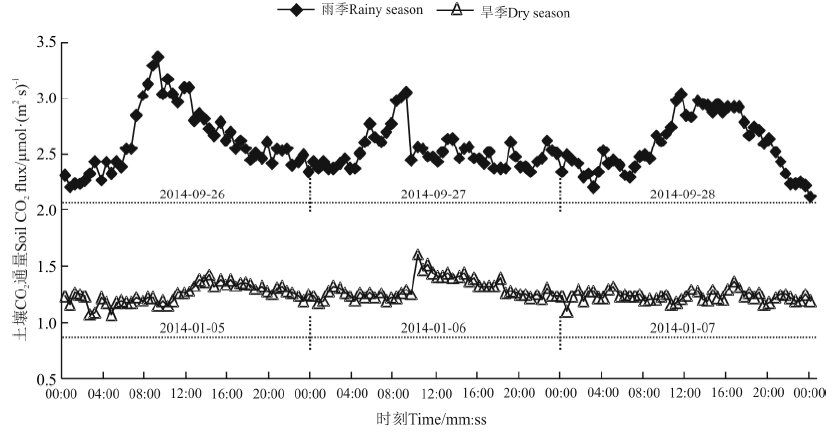 | 图1 海口城市草坪绿地土壤CO2通量日变化特征Fig. 1 Diurnal variations of soil respiration rate at a urban lawn in Haikou |
雨季日平均土壤CO2通量明显高于旱季。旱季(1月5日-1月7日)土壤CO2通量日均值1.27 μ mol· (m2· s)-1; 雨季(9月26日-9月28日)土壤CO2通量日均值2.58 μ mol· (m2· s)-1; 雨季土壤CO2通量日均值约为旱季的两倍。原因在于雨季土壤温度较高, 水分也较充足, 植物根系呼吸作用增强及土壤微生物活性较高的缘故; 而旱季则相反。
2.1.2 草坪绿地土壤CO2通量年变化特征 草坪绿地年内土壤CO2通量变化过程呈雨季高于旱季的不规则单峰曲线(图2)。年内不同月份土壤CO2通量的日平均值波动较大, 最低值出现在12月, 为0.86 μ mol· (m2· s)-1; 峰值出现在6月份, 为4.27 μ mol· (m2· s)-1, 峰值约为峰谷的5倍。随着月均温度的下降, 土壤CO2通量呈现逐渐减小趋势; 由于土壤CO2通量还受降水、土壤性状等多种环境因子的影响, 不同月份的土壤CO2通量波动较大; 同时, 雨季昼夜温差、土壤性状等变化均大于旱季, 致使雨季土壤CO2通量变幅较旱季明显。
 | 图2 海口城市草坪绿地土壤CO2通量年内变化特征Fig. 2 The monthly variations of soil respiration rate at a urban lawn in Haikou |
2.2.1 土壤CO2通量与大气温度之间的关系 城市草坪绿地土壤CO2通量随着大气温度的上升而增加, 可用指数回归模型表示(图3)。草坪绿地月均土壤CO2通量(y)与月均大气温度(x)之间的回归分析结果为y=0.309e0.078x(P=0.000, R2=0.823), 二者极显著相关。
 | 图3 海口草坪绿地土壤CO2通量与大气温度和土壤温度的指数回归Fig. 3 Exponential regressions between soil respiration rate and temperature of air and soil |
2.2.2 草坪绿地土壤CO2通量与土壤温度(5 cm)之间的关系 指数模型较好地反映土壤CO2通量与5 cm土壤温度之间的关系(图3)。草地每0.5 h的土壤CO2通量(y)与土壤温度(x)之间回归分析结果为y=0.248e0.083x (P=0.000, R2= 0.797, N=300), 二者极显著相关。
Q10是衡量土壤CO2释放对温度敏感性的重要指标, Q10越高表明土壤呼吸速率对温度的依赖性越大。海口热带草地全年土壤温度为16.2~33.8 ℃, 在土壤生物正常活动的范围(< 35 ℃)之内。旱季(日最高土壤温度< 30 ℃)Q10约为2.48, 雨季(日最高土壤温度> 30 ℃)Q10为1.44, 全年Q10为2.29。
2.2.3 草坪绿地土壤CO2通量的环境因子影响比较 为研究各环境因子对土壤CO2通量的影响, 利用SPSS 21.0对采集的数据进行统计分析(每月选取1 d, 07:00-19:00, 每0.5 h采集一个平均值, 共计300组数据)。土壤CO2通量与土壤温度、大气温度和土壤体积含水量的Pearson相关系数分别为0.845、0.877和-0.112, 双侧检验显著性水平分别为0.000、0.000和0.054。可见, 土壤CO2通量与大气温度、5 cm深土壤温度之间极显著相关, 而与土壤体积含水量相关性不显著。偏相关分析是剔除了其它相关因子的影响, 能够真实地反映各环境因子与土壤CO2通量之间关系; 结果表明, 土壤CO2通量与大气温度呈极显著正相关, 与5 cm深土壤温度呈显著正相关, 而与0-10 cm土壤体积含水量相关性不显著(表2)。
温度之间呈极显著的相关, 而与土壤体积含水量相关性不显著。偏相关分析是剔除了其他相关因子的影响, 能够真实地反映各环境因子与土壤CO2通量之间关系; 结果表明, 土壤CO2通量与大气温度呈极显著正相关, 与5 cm深土壤温度呈显著正相关, 而与0-10 cm土壤体积含水量相关性不显著(表2)。
| 表2 土壤CO2通量与影响因子的Pearson相关系数与偏相关系数(N=300) Table 2 Pearson correlation and partial correlation coefficient between soil respiration rate and its influence factors (N=300) |
2.2.4 大气降水对土壤CO2通量的影响 与晴天相比, 雨天草地土壤CO2通量变化过程更复杂, 变幅也显著增大(图4)。2014年9月7日早晨天阴, 10:00左右小雨, 14:00左右降雨强度加大, 一直持续到晚上22:00左右, 降雨总量达到100 mm。土壤水分随着降雨强度增加逐渐增多, 从20.4%增加到43.3%, 而土壤温度从29.2 ℃逐渐降到27.2 ℃。与此对应的是土壤CO2通量不断升高, 到14:30达到最高值, 由降雨前的3.27升高到20.76 μ mol· (m2· s)-1, 土壤呼吸速率增加了6倍多, 对应的土壤体积含水量、土壤温度分别为35.7%、28.7 ℃。随着降雨的持续, 土壤CO2通量逐步降低, 到19:00左右降至1.43 μ mol· (m2· s)-1, 降幅93%, 而表层0-10 cm处的土壤体积含水量上升到43.3%, 土温下降至27.2 ℃。可能因为持续降水入渗对土壤空气的挤出作用, 使土壤呼吸急剧增加; 而当降雨超过一定阈值, 土壤含水量升高甚至饱和, 降水对土壤呼吸产生抑制作用。
 | 图4 9月7日降水过程土壤CO2通量、土壤温度、土壤水分的变化Fig. 4 Variations of soil respiration rate, soil temperature and soil volumetric water content during a rainfall process in 7th September |
热带草地土壤CO2通量日均值介于0.64 ~4.99 μ mol· (m2· s)-1, 在各种陆地生态系统类型中处于较高水平。热带草地土壤CO2通量值不仅高于亚热带城郊草坪绿地[8], 也高于有机碳含量较多的高山草甸土壤CO2通量值[17]; 但其年内最大值比Michael等[18]在安大略湖北部测得的草地生态系统年内最大值要低[5.70 μ mol· (m2· s)-1)], 也略低于温带苜蓿(Medicago sativa)草地土壤呼吸[19]。土壤水分充足, 土壤呼吸随温度升高而增加; 土壤水分不足, 干旱胁迫降低土壤呼吸, 而灌溉则会显著增加旱季土壤呼吸[20]。尽管海口草地的土壤有机质含量较低, 但因地处热带, 光照强、温度高, 沙壤土透气性好, 土壤含水量较高, 故土壤呼吸较为旺盛。随着城市用地以及城市绿地面积快速扩张, 采取措施降低城市绿地土壤CO2通量值, 对缓解城市热岛效应以及应对气候变化具有重要意义, 因而需要给予更多关注。
热带海口城市草坪绿地全年Q10为2.29, 高于Bekku等[21]的研究结果(热带Q10为2.1), 也略高于同纬度的季节雨林和橡胶林土壤呼吸的Q10值(2.16和2.18)[22], 这可能与本研究区雨季降水较多, 旱季喷灌式人工浇水, 全年土壤水分含量较高有关。已有研究表明, 土壤呼吸对温度的敏感性随土壤水分的增加而增大, 即Q10随着土壤水分的升高而上升[23, 24, 25]。
较小强度降雨对土壤CO2通量变化较小; 随着降雨强度加大, 持续时间延长, 土壤CO2通量急剧增加[26]; 而当土壤水分含量较高时, 降雨则抑制土壤CO2释放速率[27, 28, 29]。土壤呼吸对降雨的响应是一个尚未十分清楚的生物地球化学过程, 降雨可能会增加或抑制土壤CO2通量。因此, 需要开展不同时间尺度上土壤CO2通量与土壤性状、土壤微生物、大气环境等多要素综合研究, 才能区别降雨入渗带来的置换效应与降雨改变底物供应的激发效应, 才能进一步阐明土壤呼吸对降雨的响应机制。
1)热带城市草地土壤CO2通量日、年变化多呈单峰曲线。土壤CO2通量日峰值一般出现在09:00-16:00, 雨季土壤CO2通量到达峰值时间较旱季要早; 6月份日均土壤CO2通量最高, 12月份最低。
2)温度是热带城市绿地土壤CO2通量较高的主要原因及其变化的关键影响因子。土壤CO2通量变化与温度呈显著的正相关关系, 而与土壤体积含水量之间的相关性不显著。
3)降雨对热带城市绿地土壤CO2通量存在挤出或抑制作用。强度较小的降雨对土壤CO2通量变化影响较小, 强度较大的降雨可导致土壤CO2通量短时间大幅增加, 而持续降雨则使土壤CO2通量受到抑制。
The authors have declared that no competing interests exist.
| [1] |
|
| [2] |
|
| [3] |
|
| [4] |
|
| [5] |
|
| [6] |
|
| [7] |
|
| [8] |
|
| [9] |
|
| [10] |
|
| [11] |
|
| [12] |
|
| [13] |
|
| [14] |
|
| [15] |
|
| [16] |
|
| [17] |
|
| [18] |
|
| [19] |
|
| [20] |
|
| [21] |
|
| [22] |
|
| [23] |
|
| [24] |
|
| [25] |
|
| [26] |
|
| [27] |
|
| [28] |
|
| [29] |
|