
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
镉胁迫对两种狼尾草不同生育时期生长性能、光合作用特征及镉吸收转运差异的影响
[刘大林1  , 孙启鑫
, 孙启鑫1 , 邵将1 , 陈鸣晖1 , 张华2 ]
, 孙启鑫, 邵将|
|


第一作者:孙启鑫(1990-),男,山东兰陵人,在读硕士生,主要从事草学研究。E-mail:[email protected]
采用盆栽法研究在不同浓度(0、5、25、50、75、100 mg·kg-1)镉胁迫下美洲狼尾草(ML, Pennisetum americanum)和杂交狼尾草(ZL, P. americanum×P. purpureum)不同生育期的生长指标、光合作用特征,并比较两者耐镉性和对镉吸收转运差异。结果表明,1)相同生育期下,随Cd浓度的增加两种狼尾草株高先升高后降低,叶宽没有显著性变化,鲜重和干重逐渐降低;2)同一生育期,ML的叶绿素a和叶绿素b含量均高于ZL;低Cd浓度(5 mg·kg-1),对植株的光合指标影响不大,但Cd浓度增加后,各处理与对照相比显著下降( P<0.05),高浓度Cd处理对ZL光合作用影响要大于ML;3)ML生长性能受Cd胁迫影响比ZL要小,但ZL富集系数高于ML,ML耐镉性更好,而ZL修复镉污染土壤的能力更强;4)根系是主要富集部位并且镉从根系往地上部位转移系数较低,3个生育期中幼苗期植株中Cd含量最高。
The growth, photosynthetic indices of Pennisetum americanum(ML) and P. americanum×P. purpureum(ZL) under different cadmium(0,5,25,50,75,100 mg·kg-1)were investigated by pot experiment and make a comparison between the two cadmium tolerance and the difference on the absorption and transport of cadmium. Results showed that: 1) the plant height decreased after increasing Cd concentration, fresh weight and dry weight decreased while leaf width changed no significant with the Cd concentration increased at the same growth stage; the indicators above increased gradually with the extension of the growth period at the same Cd concentration, however leaf width and fresh weight of ZL showed bigger; 2) At the same growth stage, Chla and Chlb content in ML were more than ZL. When Cd concentration at 5 mg·kg-1, it showed little effects on the net photosynthetic rate, transpiration rate, intercellular carbon dioxide concentration, but with the Cd concentration increased each treatment became significantly decreased compared with the control( P<0.05), which indicated that high concentrations of Cd treatment had significant effect on the photosynthesis of Pennisetum especially ZL. 3) Growth performance of American pennisetum was better than hybrid pennisetum, while the bioconcentration factor was less, indicating that the former had better tolerance to Cd, and the latter was more suitable for the remediation of cadmium contaminated soil. 4) the root system is the main bioconcentration site and Cd in root part transfered little to the overground part, in three growth stages,the content of Cd was the highest in seedling stage, which showed the seedling stage was when the plant was the most susceptible to Cd stress. ZL had a better effect than ML on the remediation of cadmium contaminated soil. If the soil under low Cd concentrations(<1.0 mg·kg-1), pennisetum still can be used as edible forage.
近年来, 随着工业化快速发展, 土壤重金属污染现在已成为人类面临的重大生态环境问题。据统计, 全世界每年向环境中释放的镉在3万t左右, 其中82%~94%的镉会进入到土壤中。我国受重金属污染的耕地约占耕地总面积的1/5, 约为2 000万hm2, 其中镉污染耕地占1 133万hm2[1]。修复土壤污染的方法有物理客土法、化学吸附法、微生物吸收法和植物修复法, 而对于大面积污染土壤, 物理、化学和微生物方法都存在相应的局限性, 难以实施。利用植物的富集作用吸收土壤中过量的重金属的植物修复技术因成本低、不破坏土壤、易实施等优点受到重点关注[2]。镉胁迫会引起植株表面中毒症状[3], 如茎叶变黄[4]、叶坏死和根褐化[5], 同时也会对光合作用[6]、酶活性[7]等生理生化进程造成不利影响, 而且对水稻(Oryza sativa)[8]、小麦(Triticum aestivum)[9]等农作物籽粒也会带来污染导致无法食用。近年来, 有关草坪草、豆科牧草对镉污染土壤修复已有较多报道[10, 11], 草坪草可以适用于非农耕区种植可以达到绿化、保持水土等作用而在农耕区会造成资源的浪费; 豆科牧草多为直根系, 对水分过多较为敏感, 在南方湿度较大的地区会引起根的腐烂[12], 是否能寻找到既不造成耕作土壤的浪费又适应南方气候的富集植物成为一个难点。美洲狼尾草(Pennisetum americanum)和杂交狼尾草(P.americanum× P. purpureum)是禾本科狼尾草属重要牧草, 狼尾草属于热带、亚热带草本植物, 喜南方高温湿润的气候条件, 植株可高达3 m, 茎叶繁茂, 须根发达, 具有生长速度快、抗旱耐涝抗病虫等优点, 同时也是优质牧草、能源草和观赏草。狼尾草主要种植于长江流域及淮河以南区域[13], 高大的生物量、广泛的用途、良好的生态习性使其具有更大种植优势, 而且在耕作区种植具有良好的经济效益, 以上特点使其具有修复植物可能性。因此, 本研究选用二倍体美洲狼尾草和三倍体杂交狼尾草为材料, 研究镉胁迫对这两种狼尾草的生长性能、光合特性的影响及这两种牧草吸收土壤中镉的效果, 为南方受镉污染土地修复和牧草利用提供依据。
试验于5月中旬在扬州大学教科园区进行, 播种基质为黏土和黄沙的混合物, 按1:3的比例配制。土壤pH为6.7, 碱解氮含量73.6 mg· kg-1, 全磷含量0.61 g· kg-1, 全钾含量5.3 g· kg-1, 有机质含量为11.3 g· kg-1, 全氮含量1.5 g· kg-1, 速效磷含量为34.3 mg· kg-1, 速效钾含量86.2 mg· kg-1, 土壤镉全量0.05 mg· kg-1 (HNO3-HC1O4-HF提取, ICP-MS测定[8])。试验材料为美洲狼尾草(ML)和杂交狼尾草(ZL), 种子均由江苏省农科院畜牧研究所提供。试验中重金属Cd 的添加形式为分析纯CdCl2· 2.5H2O, 由国药集团化学试剂有限公司生产。
因为我国南方部分农田[14]和矿区[15]土壤中Cd浓度最大值基本小于100 mg· kg-1, 平均值在2~28 mg· kg-1, 所以设定最大Cd浓度为100 mg· kg-1, 在进行正式试验之前, 先做了土壤Cd胁迫预试验, Cd浓度设定为0、2、5、10、20、30、40、50、60、70、80、90、100 mg· kg-1, 发现2 mg· kg-1虽然是严重污染程度, 但与无Cd对照生长情况无差异, 而5 mg· kg-1时出现差异, 其它浓度也表现出差异, 所以本研究设定6个Cd处理浓度(土壤中Cd浓度、以CdCl2· 2.5H2O溶液形式加入), 分别为0、5、25、50、75、100 mg· kg-1, 标记为Cd0、Cd5、Cd25、Cd50、Cd75、Cd100, 以Cd0为对照, 加入等量蒸馏水。试验为盆栽, 每盆(直径30 cm, 高25 cm)装基质土8 kg, 基肥为尿素, 按高产田水平施用(5 g· 盆-1)。于播种前两周将基质土、基肥和镉试剂混匀后装盆, 每个处理3次重复, 试验期间根据土壤情况定量浇水。待长势稳定后每盆留取15株长势相近植株, 3个生育时期分别选取5株进行测定。
1.2.1 株高、叶宽测定 每个生育期各处理随机取10株长势相近完整的狼尾草, 用直尺测定植株基部到顶部的距离为植株自然高度(cm), 每个处理3次重复; 选取从上到下第3片完全展开的完整叶片, 用游标卡尺测量叶片最宽处为叶宽(cm), 每个处理3次重复。
1.2.2 植株鲜重、干重测定 每个生育期各处理随机取若干株完整的狼尾草, 将根系洗净, 滤纸吸干根系表面的水分后, 用天平称重。计算单株的平均重量为鲜重(g· 株-1), 每个处理3次重复; 称完后将上述植株放入干燥箱105 ℃条件下, 杀青0.5 h, 并在75 ℃下烘干至恒重, 计算单株的平均重量为干重(g· 株-1), 每个处理3次重复。
1.2.3 光合指标测定 叶片气体交换参数:抽穗期在自然空气条件下, 利用美国LI-COR公司产 LI-6400系列便携式光合仪, 选取健康成熟功能叶(从分蘖外往里数第2或第3片展开叶片中部较健康、无折痕的部位), 测定时间为09:00-11:00, 测定出叶片净光合速率[Pn, μ mol· (m2· s)-1]、蒸腾速率[Tr, mmol· (m2· s)-1]、气孔导度[Gs, mmol· (m2· s)-1]、细胞间CO2浓度(Ci, μ mol· mmoL-1), 计算水分利用效率(WUE=Pn/Tr, %), 测定重复5次, 设定光照强度为1 500 lx、温度为25 ℃时的测定值进行分析。
叶绿素a和叶绿素b含量[16]:每个生育期称取0.20 g新鲜叶片(吸干表面水分), 研磨至匀浆变白, 过滤到25 mL棕色容量瓶, 用95%乙醇定容。室温下避光静置; 以95%乙醇为空白在665、649 nm下测定吸光值。根据下列公式计算:
Chla=13.95A665-6.88A649;
叶绿素a含量=Chla· V/W;
Chlb=24.96 A649-7.32 A665;
叶绿素b含量=Chb· V/W。
式中:Chla为叶绿素a的浓度(mg· L-1); Chlb为叶绿素b的浓度(mg· L-1); A665、A649分别表示665和649 nm下吸光值; V为提取后体积(L); W为叶片鲜重(g)。
1.2.4 镉含量测定 植物样测定:每个生育期, 每盆选取长势基本一致的狼尾草植株, 洗净, 将地上部分与根系分离, 105 ℃条件下, 杀青0.5 h, 并在80 ℃下烘干至恒重。烘干样品粉碎后过0.25 mm筛, 各处理称取0.20 g粉样于消煮管中, 然后加入10 mL HNO3-HCLO4(4:1)混合酸, 放置过夜后, 置于远红外消煮炉上进行消化, 待白烟冒尽, 溶液呈清凉无色, 停止加热冷却至室温, 转移至25 mL容量瓶中, 加去离子水定容, 摇匀后得试样溶液, 同时做试剂空白对照。用电感耦合等离子体质谱仪(ICP-MS)测定样品中的Cd含量。富集系数(bioconcenrration factor, BCF)和转运系数(translocation factor, TF)公式[17]如下:
BCF=植物地上部分Cd含量/土壤中总Cd含量;
TF=植物地上部分Cd含量/根中Cd含量。
采用SPSS 17.0软件对所测数据统计分析, 用平均值和标准差表示测定结果, 用Duncan法对各测定数据进行多重比较; 采用Excel 2010制图。
2.1.1 叶宽 三因素分析得出, 品种、浓度、生育时期、生育时期× 品种对植株叶宽都有显著影响(P< 0.05)。幼苗期相同浓度处理下, ZL叶宽显著高于ML(P< 0.05), ZL的叶宽随着Cd浓度的增加与对照组之间均没有显著差异(P> 0.05), ML在25和100 mg· kg-1Cd浓度时显著高于对照。分蘖期和抽穗期相同浓度处理下ML和ZL叶宽之间没有显著差异, 随着Cd浓度的增加, ML的叶宽在分蘖期和抽穗期与对照均差异不显著, ZL在抽穗期所有Cd处理与对照也无显著差异, 但在分蘖期, 75 mg· kg-1Cd浓度处理显著高于对照(图1)。
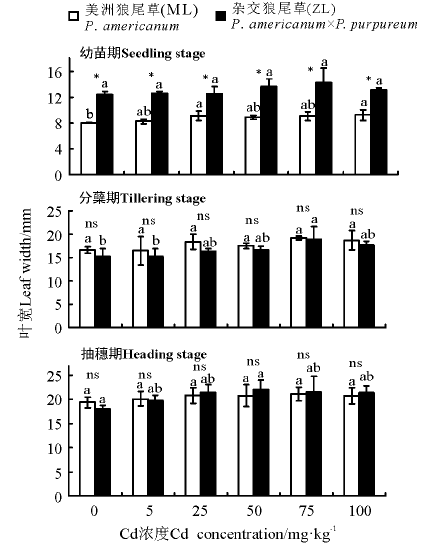 | 图1 Cd胁迫在不同生育时期对狼尾草叶宽的影响注:同一品种不同小写字母表示不同Cd浓度处理间差异显著(P< 0.05), ns表示同一浓度不同品种间差异不显著(P> 0.05), * 表示同一浓度不同品种间差异显著(P< 0.05), 下同。Fig.1 Effects of Cd stress on the leaf width of Pennisetum during different growth stagesNote: Different lowercase letters indicate significant difference in different concentrations of the same species at the 0.05 level, ns indicates no significant difference in different species of the same concentration(P> 0.05), * indicates significant difference in different species of the same concentration at the 0.05 level, similarly for each of the following figures. |
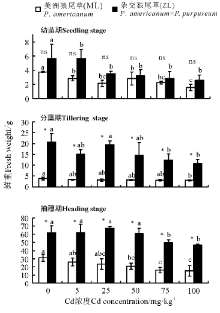
2.1.2 株高 三因素分析得出, 浓度、品种、生育时期以及生育时期× 品种对植株株高都有显著影响(P< 0.05)。3个生育时期ML株高均高于ZL, 且在分蘖期和抽穗期两者间呈显著差异(P< 0.05), 幼苗期除对照外两者均无显著差异(P> 0.05)。随着Cd浓度增加两种牧草的株高大体上呈降低趋势, 幼苗期和分蘖期ML在Cd浓度为100 mg· kg-1时均显著低于对照, ZL各处理浓度与对照相比均不显著(P> 0.05); 抽穗期ZL的100 mg· kg-1处理显著低于对照(图2)。
 | 图2 Cd胁迫对不同生育时期下狼尾草株高的影响Fig.2 Effects of Cd stress on the height of Pennisetum during different growth stages |
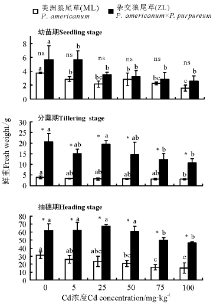
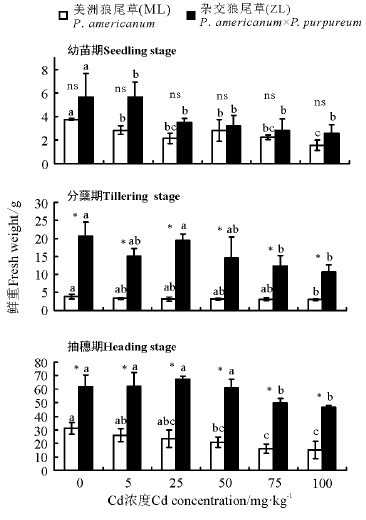
2.1.3 鲜重 三因素分析得出, 品种、浓度、生育时期、生育时期× 品种、生育时期× 浓度对植株的鲜重都有显著影响(P< 0.05)。3个生育时期相同浓度下ZL鲜重均大于ML, 幼苗期ML和ZL各处理浓度均显著低于对照且两者之间无显著差异(P> 0.05); 分蘖期ZL鲜重均显著高于ML, 且ML在Cd浓度为100 mg· kg-1时鲜重显著小于对照, ZL在Cd浓度为75和100 mg· kg-1时显著小于对照; 抽穗期ML鲜重随着Cd浓度的增加逐渐降低, 而ZL则随Cd浓度增加先上升后降低, 两者在Cd浓度为75、100 mg· kg-1时都显著小于对照(P< 0.05)(图3)。
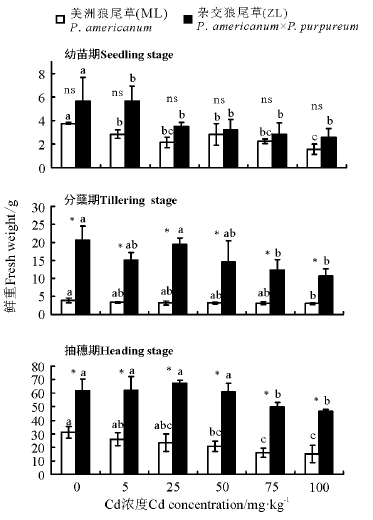 | 图3 Cd胁迫对不同生育时期狼尾草鲜重的影响Fig.3 Effects of Cd stress on the fresh weight of Pennisetum during different growth stages |
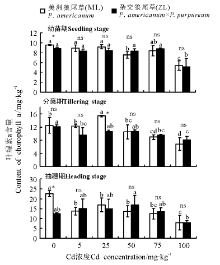
2.2.1 叶绿素a含量 三因素分析得出, 浓度、生育时期、品种× 浓度对植株叶片的叶绿素a含量都有显著影响(P< 0.05)。除分蘖期25 mg· kg-1Cd处理外, 3个生育时期两种狼尾草叶绿素a含量在相同Cd胁迫浓度下均无显著差异(P> 0.05)。幼苗期和分蘖期ML和ZL在Cd浓度为100 mg· kg-1时叶绿素a含量显著低于对照组(P< 0.05); 抽穗期随着Cd浓度的增加, ML对照组显著高于其它处理浓度(P< 0.05), 而ZL各个浓度处理与对照间均不存在显著差异(P> 0.05)(图4)。
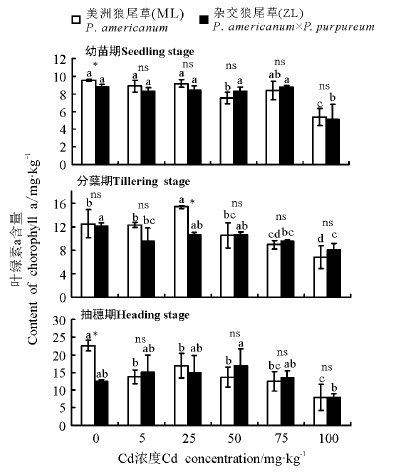 | 图4 狼尾草在三个生育时期的叶绿素a含量Fig.4 Content of chlorophyll a in different Pennisetum during different growth stages |
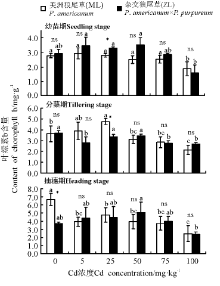
2.2.2 叶绿素b含量 三因素分析得出, 浓度、生育时期、品种× 浓度对植株叶片叶绿素b含量都有显著影响(P< 0.05)。3个生育时期两种狼尾草总体没有显著差异, 幼苗期相同处理浓度下, 随着Cd浓度的增加, ML叶绿素b含量整体呈下降的趋势, 在浓度达到100 mg· kg-1时, 叶绿素b含量显著低于对照, ZL各个处理组与对照之间无显著差异(P> 0.05); 分蘖期ML随着Cd浓度增加, 总体呈先升后降趋势, ZL叶绿素b含量则总体呈逐渐下降趋势, 且在5、75、100 mg· kg-1浓度时显著低于对照(P< 0.05); 抽穗期随着Cd浓度的增加, ML对照组叶绿素b含量显著高于其它浓度处理, 而ZL各Cd浓度处理与对照间无显著差异(P> 0.05)(图5)。
 | 图5 狼尾草在三个生育时期的叶绿素b含量Fig.5 Content of chlorophyll b in different Pennisetum during different growth stages |
2.2.3 光合参数 随着Cd浓度的增大, 两种牧草叶片的Pn、Tr、Gs逐渐呈下降趋势, Ci总体呈先升后降趋势, 但也存在差异(表1)。ML的Pn在Cd浓度为25 mg· kg-1时开始起显著低于对照(P< 0.05); 75和100 mg· kg-1Cd处理的Tr显著小于对照(P< 0.05); 各浓度处理的Gs都显著低于对照; 25、50和100 mg· kg-1 Cd浓度下的Ci都显著高于对照; WUE各浓度处理与对照间均不显著(P> 0.05)。ZL的25、50、75和100 mg· kg-1 Cd处理下Pn、Tr均显著小于对照, 且随着Cd浓度的增加Pn、Tr持续降低; 各浓度处理的Gs均显著小于对照; Ci无明显变化规律, 100 mg· kg-1 Cd处理著高于对照; WUE在75和100 mg· kg-1处理下显著小于对照。
| 表1 分蘖期Cd胁迫对狼尾草净光合速率、蒸腾速率、气孔导度、胞间二氧化碳浓度和水分利用效率的影响 Table 1 Effects of Cd stress on the net photosynthetic rate(Pn), transpiration rate(Tr), stomatal conductance(Gs), CO2 concentration(Ci) and water use efficiency(WUE) of Pennisetum in tillering stage |
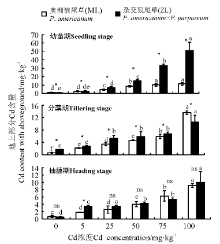
2.3.1 Cd胁迫对狼尾草地上部分Cd含量的影响 三因素分析得出, 品种、浓度、生育时期、品种× 浓度、生育时期× 品种、生育时期× 浓度和生育时期× 品种× 浓度都对植株地上部分Cd含量有显著影响(P< 0.05)。3个生育时期中随着Cd浓度的增加, 两种狼尾草地上部分Cd含量逐渐增加, Cd浓度高于5 mg· kg-1的处理均显著高于对照。幼苗期ZL在同一浓度处理下总体均显著高于ML(P< 0.05); 分蘖期Cd浓度为0~75 mg· kg-1时ZL显著高于ML, 100 mg· kg-1时ML显著高于ZL。当Cd浓度为100 mg· kg-1时地上部分Cd含量都显著高于其它浓度处理和对照; 抽穗期各浓度处理下两种狼尾草均显著高于对照, 同一浓度总体上ZL大于ML, 除5 mg· kg-1处理下ZL显著高于ML外, 其它处理下均无显著差异(P> 0.05)(图6)。
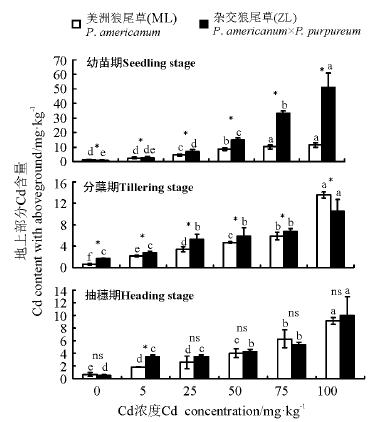 | 图6 3个生育时期狼尾草地上部分Cd含量Fig.6 Cd content with aboveground of Pennisetum during different growth stages |
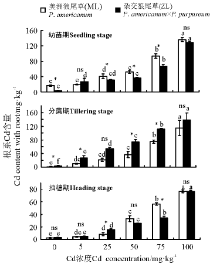
2.3.2 Cd胁迫对狼尾草根系Cd含量的影响 三因素分析得出, 浓度、品种、生育时期、生育时期× 品种、生育时期× 浓度、生育时期× 品种× 浓度对植株根系Cd含量均有显著性影响(P< 0.05)。3个生育时期, Cd浓度为50、75和100 mg· kg-1时两种牧草均显著高于对照(P< 0.05), 且随着Cd浓度的增加根系中Cd含量逐渐增加; 除5 mg· kg-1 Cd处理外, 幼苗期相同浓度下ML根系Cd含量均高于ZL; 分蘖期相同浓度下ML根系Cd含量均低于ZL; 抽穗期25 mg· kg-1 时ZL显著大于ML, 75 mg· kg-1时ML显著大于ZL(图7)。
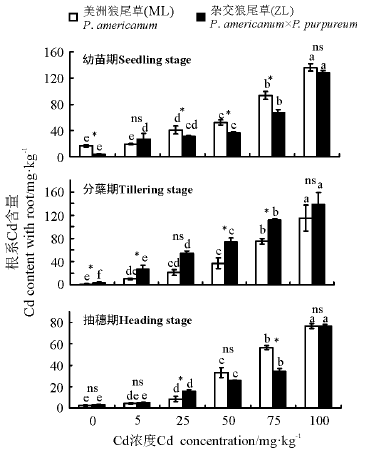 | 图7 3个生育时期狼尾草根系Cd含量Fig.7 Cd content with roots of Pennisetum during different growth stages |
2.3.3 Cd胁迫对狼尾草不同生育期转移系数和富集系数的影响 两种狼尾草在抽穗期的转移系数和3个生育期的富集系数总体随着Cd浓度的增大呈降低趋势(表2)。ML在分蘖期和抽穗期时土壤Cd浓度为5 mg· kg-1时的转移系数最大, 75 mg· kg-1时最低, 幼苗期以Cd浓度为50 mg· kg-1时转移系数最大, 100 mg· kg-1最小; 抽穗期总体高于幼苗期和分蘖期; 转运系数总体为7.88%~44.25%。富集系数方面, ML在3个生育时期均以Cd浓度为5 mg· kg-1处理显著高于其它浓度处理(P< 0.05), 而随着生育时期的延长略显降低, 其范围为7.86%~46.61%。ZL转移系数与ML表现类似, 但幼苗期不同, 随着Cd浓度增加呈先升高后降低趋势, 当Cd浓度为75 mg· kg-1时表现值最大; 3个生育时期的转运系数范围为6.09%~76.36%。ZL的Cd浓度为5 mg· kg-1时处理富集系数显著高于其它浓度处理, 但两种牧草的富集系数有所不同, 同一生育时期内, ZL富集系数通常高于ML, 范围为7.17%~69.56%。
| 表2 Cd胁迫对狼尾草不同生育期转移系数和富集系数的影响 Table 2 Effects of Cd stress on the transfer factor and bioconcentration factor of Pennisetum during different growth stages |
Cd是植物的非必需元素, 一定浓度会严重阻碍植物生长甚至死亡。有研究发现, 低浓度的Cd2+能够促进野燕麦(Avena fatua)幼苗株高和地上、地下部分干、鲜重量的积累, 高浓度Cd2+能抑制野燕麦幼苗生长[18]。紫花苜蓿(Medicago sativa)的株高随镉浓度增加而降低, 鲜重则先升高后降低[19]。本研究结果显示, 两种狼尾草的株高随Cd浓度增加先稍微增高后再逐渐下降, 这也说明了低浓度的Cd对植株生长有一定的刺激作用; 而叶宽却没有表现出明显的变化, 这可能是茎叶生长内在原因不同:茎中含有较多生长素, Cd可能抑制生长素的合成或运输, Cd会对叶片细胞中类囊体等造成损害[20], 但应该没有破坏细胞壁; 鲜重和干重的逐渐降低也反应出高浓度Cd加重了对植株生长的抑制效果。尽管狼尾草株高、叶宽受Cd胁迫导致平均鲜重下降, 但Cd浓度为100 mg· kg-1时仍随着生育期延长而增大, 可见狼尾草对Cd胁迫有良好的耐受性。Cd对两种狼尾草抑制作用为株高> 叶宽, 杂交狼尾草> 美洲狼尾草, 这说明叶比茎耐受性大, 美洲狼尾草对Cd的耐受性强于杂交狼尾草。
叶绿素是光合作用的的最重要色素, 直接反映光合效率大小, 高等植物中叶绿素由叶绿素a和叶绿素b组成。当植物受到重金属离子胁迫时, 叶绿素合成和光合作用一般会受到抑制, 使叶绿素总量和光合效率降低。一般认为光合速率降低原因有:Gs与Ci相同变化时为气孔因素限制; Gs与Ci相异变化时为非气孔因素限制[21]。有研究发现, 镉胁迫下菜豆叶片中光合色素含量随着Cd浓度的增加和时间的延长呈下降趋势[22]。但本研究结果显示, 总体上叶绿素a和叶绿素b含量随Cd浓度增加逐渐下降, 但有些生育期中呈先升后降趋势, 这与刘周莉等[23]研究结果相似。也有研究表明, Cd浓度的增加抑制匍匐剪股颖(Agrostis stolonifera)叶绿素含量[24]。这可能是植株体内摄入过多的Cd, 造成叶绿素酶活性比例失调, 加快叶绿素分解, 从而导致含量下降。随着生育期的延长, 两种叶绿素的含量也在增加, 但抽穗期的最高浓度处理都显著低于幼苗期的低浓度(5 mg· kg-1), 说明较低浓度Cd处理对狼尾草的叶绿素含量较小, 浓度比时间的胁迫影响更强些。叶绿素a是光反应的中心色素, 叶绿素b则捕获光子, 叶绿素a和b含量使光反应受阻, 气孔导度下降和胞间二氧化碳浓度变化使得净光合速率下降。
陈良等[25]研究表明, Cd胁迫下, 两种菊芋(Helianthus tuberosus)幼苗的Pn、Gs、Tr值均明显下降, Ci较稳定, 能通过增加叶片的类胡萝卜素含量缓解Cd的抑制作用。本研究结果显示, 两种狼尾草的Pn、Gs、Tr值亦均明显下降, 美洲狼尾草的Ci值先升后降, 因此其光合速率的降低是由气孔因素造成, 即叶肉细胞光合活性降低所致; 杂交狼尾草的Ci变化不规律, 处理组高于对照说明气孔因素和非气孔因素都会影响其光合速率, 这与孙璐等[26]研究结果相似。蒸腾速率降低是植物的一种自我保护反应, 减少对水分吸收同时也减少对重金属离子的吸收。水分利用率是显示植物水分吸收利用过程效率的一个指标[27], 其值逐渐降低显示出遭受Cd胁迫程度也在增大。
许多研究表明不同的植株对Cd吸收能力存在差异, 这些耐性植物能在根部富集大量重金属离子, 而限制向地上部分运输, 从而使地上部分免受伤害, 一定程度提高植株的耐性[28]。
比如大豆(Glycine max)[29]、小白菜(Brassica rapa)[30]、宝山堇菜(Viola baoshanensis)[21]等品种对Cd都能吸收, 而宝山堇菜是超富集植物, 其对Cd具有很强的吸收能力, 这是由于植物的遗传特性和自身生物特点所决定的。研究发现, 重金属Cd胁迫下, 印度芥菜( Brassica juncea)根部Cd离子含量明显高于地上部分[31]。本研究中地上部分和根系中Cd含量都随Cd浓度的增大而增大, 而根系显著高于地上部分和转运系数都小于1, 说明狼尾草根部是主要的Cd富集器官, 这表现为普通植物的特征, 并不满足超富集植物的要求, 且Cd从根部到地上的转运能力较小, 这与狼尾草具有发达的须根系和较高的株高密切相关。当土壤中Cd浓度高达100 mg· kg-1时生长状况仍良好, 所以狼尾草对镉胁迫具有良好耐性。
富集系数在土壤镉浓度为5~25 mg· kg-1时下降迅速, 随后下降缓慢, 说明低浓度狼尾草主动吸收Cd, 高浓度抑制对Cd的吸收[32]。有研究发现, 土壤中Cd含量为1.57 mg· kg-1时, 水稻富集系数为0.56[33]; 土壤镉浓度为0.28 mg· kg-1时, 小麦富集系数为0.49[34]; 土壤中Cd含量为0.35 mg· kg-1时, 苜蓿富集系数为0.93[11]。本研究中低浓度Cd处理时富集系数为0.45, 这可能是相同的科中存在某种类似的转移机制或者体内其它元素拮抗导致。
镉胁迫抑制了两种狼尾草的生长、光合作用和蒸腾作用, 随着镉浓度增加和生育期的延长, 抑制作用加强, 其中美洲狼尾草耐受性强于杂交狼尾草, 但杂交狼尾草吸收镉的能力要优于美洲狼尾草。
The authors have declared that no competing interests exist.
| [1] |
|
| [2] |
|
| [3] |
|
| [4] |
|
| [5] |
|
| [6] |
|
| [7] |
|
| [8] |
|
| [9] |
|
| [10] |
|
| [11] |
|
| [12] |
|
| [13] |
|
| [14] |
|
| [15] |
|
| [16] |
|
| [17] |
|
| [18] |
|
| [19] |
|
| [20] |
|
| [21] |
|
| [22] |
|
| [23] |
|
| [24] |
|
| [25] |
|
| [26] |
|
| [27] |
|
| [28] |
|
| [29] |
|
| [30] |
|
| [31] |
|
| [32] |
|
| [33] |
|
| [34] |
|