
{kind=link}
蒙古冰草和冰草的杂交F1代群体农艺性状的相关、通径及聚类分析
[段清清1  , 徐荣蕊
, 徐荣蕊1 , 李旭青1 , 车永和1 , 张锦鹏2 , 李秀全2 , 杨欣明2 , 刘伟华2 ]
, 徐荣蕊, 张锦鹏]
|
|


第一作者:段清清(1990-),女,河南南阳人,在读硕士生,主要从事作物遗传育种研究。E-mail:1589355 [email protected]
通过探究冰草属( Agropyron)植物的主要农艺性状间的关系,选育出优良冰草( A. cristatum)种质资源为提高小麦( Triticum aestivum)远缘杂交育种效率提供理论依据。本研究以蒙古冰草( A. mongolicum)(母本)和二倍体冰草(父本)进行杂交获得的F1代群体为材料,对该群体116个单株的9个主要农艺性状进行了调查分析。结果表明,116个单株在所有农艺性状上均表现出明显的遗传变异(11.2%~40.13%),小穗粒数、穗粒数的变异系数最大,分别为30.56%、40.13%;相关及通径分析表明,对其重要性状之一千粒重影响最大的是每小穗小花数( r=0 .168, P<0 .05),直接通径系数为0 .364,其余性状依次为穗长( r=0 .235, P<0 .05)、小穗粒数( r=0.126);聚类分析在欧氏距离为6.50处将群体材料分成三大类群,其中第1类和第3类冰草材料都具有高秆、多枝叶的特点,可用于选育优质牧草,第1类冰草材料具有多小花多穗粒等优良性状,可成为饲草和小麦遗传育种的优良种质资源。
, Xu Rong-rui, Zhang Jin-peng
To explore the relationship among nine agronomical characteristics of Agropyron. and provide the theoretical basis for improving the efficiency of wheat breeding, the F1 generation of the cross between Agropyron mongolicum (female parent) and the diploid A. cristatum (male parent) was analysed. The nine agronomic characteristics of 116 F1 plants was analysed using different methods of data analysis. The results showed that all characteristics exhibited significant genetic variation (11.2%~40.13%). The variation in grains per spikelet and grains per spike were the greatest, with a value of 30.56% and 40.13% respectively. Correlation and path analysis showed that florets per spikelet had a significant effect on thousand kernel weight with an r value of 0.168 ( P<0.05) and a direct path coefficient of 0.364. This was followed by spike length (r = 0.235, P<0.05) and kernels per spikelet (r=0.126). The 116 plants of the Agropyron F1 population were clustered into three groups. The first and third group had greater plant height and more foliage. As such, these groups could represent varieties with high-quality forage grass for breeding. The first and third group had numerous positive characteristics, including more florets per spikelet, more spike grains, among others, which represent good germplasm resources for forage grass and wheat breeding.
冰草属(Agropyron)植物为禾本科小麦族多年生异花授粉植物, 主要分布于欧亚大陆草原区, 生态幅度十分广阔[1]。其中蒙古冰草(Agropyron mongolicum)和冰草(A. cristatum)是冰草属常见的两个冰草种, 蒙古冰草又称沙芦草, 主要分布于我国的陕西、宁夏、内蒙古、新疆、甘肃、山西等省(区)的草原或沙地[1, 2], 其属于我国二级植物保护名录植物[3]; 冰草在我国东北、西北及内蒙古自治区等地有野生种分布, 抗寒、耐旱, 作为优良牧草已被栽培利用[4]。牧草是农业生产中重要的生产资料之一, 选育优质饲草对我国牧草产业的良性发展具有重大作用[5]。冰草属植物草质柔软, 适口性良好, 营养价值高, 可用作青饲、晒制干草、制作青贮或放牧, 目前作为优良的牧草广为栽培, 在农牧业生产中具有很高的经济价值[6]。此外, 它还是主要的固沙草本植物和草坪用种, 其根系庞大, 抗逆性强, 用于我国北方干旱半干旱地区改良草地, 建立栽培草地, 是优良的护坡、保持水土和绿化植被, 在保持生态平衡方面具有重要的价值[7]。同时, 冰草属植物作为小麦的重要野生近缘属之一, 其具有极强的抗旱、抗寒和耐盐碱的特性[8], 以及抗赤霉病、叶锈病等优良特性, 还兼具高千粒重、多花、多小穗、多分蘖等丰产特性, 是改良小麦的优质种质资源[9]。目前已培育出小麦(Triticum aestivum)-冰草6P附加系[10]、小麦-冰草6P染色体中间插入型易位系[11]和小麦-冰草5A/6P 易位系[12]等一系列远缘杂交材料。
农艺性状是选育植物新品种的重要标记, 有助于植物品种间表型变异及亲缘关系的分析[13]。表型多样性是遗传多样性的基础, 因此, 对于冰草这样分布广, 变异性明显的物种来讲, 通过调查农艺性状研究冰草的遗传多样性是十分有效的。例如, 从形态学角度对14个冰草种群的10个穗部性状进行了测定分析, 揭示了冰草种内不同种群间及种群内的不同个体间的遗传变异[14] ; 应用居群生物学原理对35个蒙古冰草居群的形态学性状进行了分析, 结果表明居群内存在丰富的遗传多样性[2]。农艺性状分析主要反映物种基因与环境相互作用后物种的性状表现[15], 且大多数为数量性状, 因此利用数量遗传学方法诸如相关分析、通径分析及聚类分析等进行研究, 方法简单、有效, 不仅可以揭示其遗传背景及其内在联系, 还有助于进行品种改良, 提高育种效率, 从而有效保护和合理利用这一种质资源[14]。
冰草属植物研究中已利用二倍体冰草为父本、蒙古冰草为母本杂交获得的F1代群体, 即异花授粉CP(cross-pollinated)群体为作图群体, 结合“ 拟测交” 分析策略, 采用SLAF测序技术检测到的多态性SNP分子标记构建了冰草P基因组遗传连锁图谱, 包含1 023个分子标记共7个连锁群, 图谱全长907.8 cM, 平均图距1.5 cM[16]。目前, 有关冰草属F1代群体的农艺性状分析尚未见报道, 而重要农艺性状的研究是发现其相关分子标记的基础, 对开展相关植物分子育种具有重要意义。本研究采用以冰草属典型物种蒙古冰草居群和二倍体冰草进行人工杂交获得的杂交F1代群体为试验材料, 调查冰草F1代群体各单株的主要农艺性状, 并通过对主要农艺性状进行的相关、通径及聚类分析, 重点了解各农艺性状间的相互关系及其对改良小麦重要性状之一千粒重的影响程度, 以期从中选育出优良的冰草种质资源, 为拓宽冰草的遗传基础, 提高牧草育种的选择效率提供理论依据, 同时选出冰草新材料, 为冰草的优良基因在小麦遗传改良中得到有效应用奠定基础。
试验材料由中国农业科学院作物科学研究所提供, 全部材料种在廊坊中国农业科学院国际农业高新技术产业园区。F1代群体是以二倍体冰草Z1842为父本, 蒙古冰草Z2098为母本, 采用捻穗法进行杂交获得的。本研究采用F1代群体中的116个单株材料进行分析。每份材料进行3个重复处理, 行距80 cm, 株距40 cm, 并进行田间统一种植管理。
依据《小麦种质资源描述规范和数据标准》考察农艺性状[17]。试验种植地廊坊地处中纬度地带, 属暖温带大陆性季风气候, 种植地土壤为沙壤土。2014年7月中旬进行考种, 冰草F1代群体共统计了116株冰草植株的数据。每株冰草植株分别调查株高、穗茎节间长、穗长、穗宽、每穗小穗数、每小穗小花数、小穗粒数、穗粒数、千粒重等农艺性状; 具体调查方法:先测量每一个冰草植株的株高, 然后每一株随机选取10个穗子, 调查穗长、穗宽、穗茎节长、每穗小穗数、每小穗小花数、小穗粒数和穗粒数7个农艺性状, 取平均值, 最后将冰草的穗子脱粒, 统计千粒重。
应用Excel进行数据统计, 利用SPSS 20进行遗传变异分析、相关性分析、通径分析和聚类分析。通径分析采用线性回归的方法进行[18], 并设定:X1, 株高(cm); X2穗茎节间长(cm); X3, 穗长(cm); X4, 穗宽(mm); X5, 每穗小穗数(个); X6, 每小穗小花数(个); X7, 小穗粒数(粒); X8, 穗粒数(粒); Y, 千粒重(g)。
对研究群体9个主要农艺性状分析结果(表1)表明, 116份群体材料间的主要农艺性状存在丰富的变异。株高平均为85.79 cm, 变化范围为50.00~124.00 cm, 极差值达到同一性状最小单株的两倍以上。穗茎节长平均为37.28 cm, 变化范围为27.44~46.52 cm。穗长平均为8.08 cm, 变化范围为4.80~10.84 cm。穗宽平均为9.41 mm, 变化范围为6.60~12.67 mm。每穗小穗数平均为27.45个, 变化范围为12.60~37.20个。每小穗小花数平均为7.37个, 变化范围为4.67~10.10个。小穗粒数平均为1.24个, 变化范围为0.30~2.13个。穗粒数平均为18.67个, 变化范围为0.40~43.1个。千粒重平均为2.31 g, 变化范围为1.80~3.23 g。
| 表1 冰草F1群体主要农艺性状变异分析 Table 1 Variation of Agropyron F1 populations in main agronomic characters |
研究群体各农艺性状的变异系数在11.26%~40.13%, 穗粒数的变异系数最大, 为40.13%, 其次是小穗粒数, 其变异系数为30.56%; 株高、每穗小穗数、每小穗小花数、穗宽、穗长、千粒重和穗茎节长的变异系数分别为15.02%、13.61%、13.31%、12.03%、11.85%、11.71%和11.26%, 变异较明显。
对种间F1代群体9个农艺性状的相关分析结果表明, 共有10对性状达到极显著相关(P< 0.01), 4对性状达到显著相关水平(P< 0.05)。其中, 株高与穗茎节长(r=0.359, P=0)呈极显著正相关。穗茎节长与千粒重(r=-0.190, P=0.048)呈显著的制约关系。穗长则与每穗小穗数(r=0.585, P=0)、穗茎节长(r=0.286, P=0.003)、每小穗小花数(r=0.413, P=0) 呈极显著的正相关, 并与千粒重(r=0.235, P=0.014) 呈显著的正相关, 却与穗宽呈极显著负相关的制约关系(r=-0.373, P=0), 与此同时, 穗宽与每穗小穗数(r=0.420, P=0)、每小穗小花数(r=0.641, P=0) 均呈极显著的正相关。这说明, 穗长与穗宽相互制约的关系也限制了冰草的小穗数和小花数, 而每穗小穗数与每小穗小花数(r=-0.588, P=0)、小穗粒数(r=-0.244, P=0.011)呈现极显著和显著负相关的制约关系, 但与穗粒数达到极显著的正相关(r=0.362, P=0), 与之存在制约关系的每小穗小花数对穗粒数(r=0.232, P=0.015)都有显著影响, 均呈现显著正相关性, 表明决定产量的要素之间存在着相互制约的关系。此外, 小穗粒数与穗粒数之间存在极显著正相关性(r=0.762, P=0), 但受限于每穗小穗数以及穗长与穗宽等农艺性状的制约。
| 表2 冰草F1群体主要农艺性状的相关性分析 Table 2 Correlation of main agronomic characters of Agropyron F1 populations |
冰草属植物是改良小麦的优质资源。千粒重是构成主要农作物产量的三因素之一, 是体现种子大小与饱满程度的一项指标, 也是田间预测产量时的重要依据[19]。因此, 针对冰草属植物千粒重进行回归分析有重要意义。以观测到的株高、穗茎节长、穗长、穗宽、每穗小穗数、每小穗小花数、小穗粒数和穗粒数共8个农艺性状为自变量, 千粒重为因变量, 进行逐步回归分析, 得到最优回归方程为Y=0.920+0.060X3+0.101X6+0.135X7, 方程中共筛选出3个因子, 其中, 穗长、每小穗小花数与千粒重的相关性达到显著水平(P< 0.05), 相关系数分别为0.235、0.168。另外, 根据回归方程的相关系数为R2=0.245, 可得剩余因子e=0.872[18, 20], 该值较大, 说明影响冰草千粒重的主要因素尚不能确定。
根据通径分析原理, 每一性状对千粒重的相关系数由该性状对千粒重的直接效应和间接效应所组成[21]。3个农艺性状对千粒重的直接通径系数有差异(表3), 对千粒重作用最大的是每小穗小花数, 直接通径系数为0.364, 其余依次为穗长、小穗粒数, 直接通径系数分别为0.212、0.188。每小穗小花数对千粒重的直接作用最大, 与直接作用相比, 其通过穗长、小穗粒数的间接作用被削弱, 间接通径系数分别为0.036 0和0.008 5。穗长、小穗粒数对千粒重的间接作用较高, 穗长通过每小穗小花数的间接通径系数为0.061 9, 小穗粒数通过每小穗小花数的间接通径系数为0.016 4, 但穗长与小穗粒数之间对千粒重的间接作用被削弱。3个影响因素对千粒重均为正向促进作用, 结合相关分析和回归分析的结果可知, 穗长、每小穗小花数是千粒重的重要影响因子, 每小穗小花数对千粒重的贡献最大。
| 表3 冰草F1 群体主要农艺性状对千粒重的通径分析 Table 3 Path analysis of main agronomic characters of Agropyron F1 populations |
根据研究群体9个主要农艺性状的考种数据, 对群体材料进行系统聚类分析, 结果表明在欧氏距离为6.50处可以将116个冰草F1代单株聚为3类(图1), 类群组成及各类群主要农艺性状的平均值如表4所示。第1类包含56个冰草F1代群体单株; 第2类包含7个冰草F1代群体单株; 第3类包含53个冰草F1代群体单株。第1类和第3类占了大多数, 第2类数量很少, 有F1-14、F1-22、F1-64、F1-97、F1-114、F1-119以及F1-128共7个冰草单株, 为矮秆、长穗、少花少粒、低千粒重型。第1类明显特征表现为植株较高, 宽穗, 多小花多小穗粒, 千粒重最高, 为中秆、宽穗、多花多粒粒型; 第3类明显特征表现为植株高度最高, 穗短而宽, 小穗数、小穗粒数较少, 小花数较多, 千粒重较高, 为高秆、短穗紧密、少穗粒型。
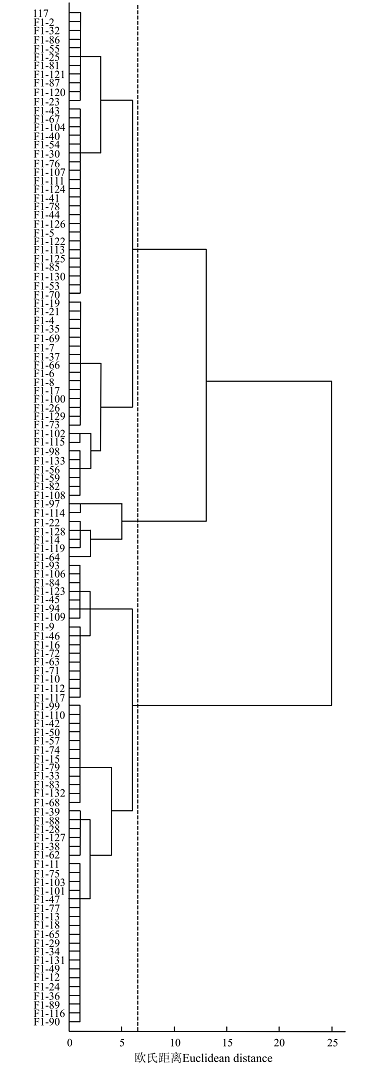 | 图1 研究群体主要农艺性状的Ward联接系统聚类图Fig. 1 Cluster analysis of main agronomic characters of the studied populations |
| 表4 冰草F1群体3个类群各农艺性状的平均值 Table 4 Means of all agronomic characters of Agropyron F1 populations in three clusters |
综合上述分析, 冰草属种间F1代群体单株在株高、穗部性状等农艺性状上存在明显差异, 性状的差异显著性各不相同。穗粒数和小穗粒数的变异系数分别达到40.13%、30.56%, 说明冰草F1群体里存在着具有多穗粒的优良性状的单株, 其遗传变异的差异远大于其它农艺性状的差异。其余农艺性状的变异系数也达到了相当高的水平(11.26%~15.02%), 其中株高的遗传变异表现最为显著, 每穗小穗数等性状在各单株间也有明显差距。可见, 冰草F1群体种质资源在多个农艺性状上差距明显, 类型广泛, 有一定的选择范围优势。这种差异可能来源于几个方面。首先, 作为亲本的蒙古冰草和冰草不同物种间遗传基础差异较大, 其杂交F1代的遗传物质有较丰富的变异; 其次, 自然环境和人工等因素也会影响冰草F1群体的生长, 促使其表型多样性的增加。种质资源的遗传多样性是育种工作的基础, 因此提高育种基础材料的遗传多样性是十分必要的[22]。冰草属植物种间杂交丰富了后代材料的遗传变异, 提高了遗传改良材料的选择利用性。
相关系数是表明各性状间的相关程度, 分析结果表明, 各个性状间既存在有促进作用的显著正相关, 又存在有制约作用的显著负相关性。在进行性状选择时, 不但要注重目标性状的选择, 还应注意到与它们存在相互制约关系的其它农艺性状。如株高、穗长、穗宽、每穗小穗数、每小穗小花数等多个农艺性状都对最终产量有影响, 当选择多穗粒的性状时, 必须综合利用其它农艺性状对产量的促进作用以及制约关系, 这将有利于提高冰草品种改良的效率, 也为其优异基因资源在小麦育种中的应用提供参考。
通径分析和多元回归分析有互补作用, 多元线性回归分析可以比较多个相关性较大的原因与结果的关系, 且结果具有预测性, 但是不能反映自变量之间的相互关系, 所以存在一定的局限性, 需要进行通径分析来加以完善[20]。通径分析着重讨论各个原因对结果影响的重要程度[22], 是研究和处理不同数量性状之间关系的一种有效方法[20]。目前在扁蓿豆(Medicago ruthenica)[20]、白羊草(Bothriochloa ischaemum)[22]和苏丹草(Sorghum sudanense)[23]等牧草上已经广泛应用。分析结果表明, 每小穗小花数、穗长和穗粒数对提高千粒重有促进作用, 但对这两个农艺性状有显著影响的穗宽和每穗小穗数之间存在复杂的联系, 其中穗长和穗宽是相互制约的关系, 每穗小穗数又与每小穗小花数是呈极显著负相关的制约关系, 因此无法单纯通过某一性状的提高来达到提高千粒重的目的。通径系数能够说明相关性状的因果关系及各性状对千粒重提高的相对重要性大小, 穗粒数与千粒重的相关性虽不显著, 但通径分析却显示穗粒数对千粒重有比较重要的作用。在可促进提高千粒重的农艺性状中, 每小穗小花数、穗粒数和穗长这3个影响因素所占比重并不大, 因此, 要选育出较高产种量的冰草材料, 还需对千粒重的影响因素的进一步分析研究。
根据9个农艺性状考种数据对冰草F1群体的116个单株进行聚类分析, 将其聚为三大类。聚类图显示冰草F1群体分化较大, 说明冰草F1群体各个材料有较大的遗传差异。其中, 第3类有最高的株高, 第1类有最多的小穗数, 并具有多花多粒的性状, 均为优良的种质资源, 这两类材料的枝繁叶茂特性可用于选育优质牧草, 不同类材料的高千粒重、多花多粒等特异性状对小麦远缘杂交育种有指导意义。目前, 利用本研究的F1群体研究人员已构建相应的分子图谱[16], 通过我们进一步形态学和农艺性状调查将有望构建重要性状的QTL分子图谱。相关冰草F1群体的分类也有待进一步研究。
The authors have declared that no competing interests exist.
| [1] |
|
| [2] |
|
| [3] |
|
| [4] |
|
| [5] |
|
| [6] |
|
| [7] |
|
| [8] |
|
| [9] |
|
| [10] |
|
| [11] |
|
| [12] |
|
| [13] |
|
| [14] |
|
| [15] |
|
| [16] |
|
| [17] |
|
| [18] |
|
| [19] |
|
| [20] |
|
| [21] |
|
| [22] |
|
| [23] |
|