

{kind=link}
{kind=link}
千日红无菌苗生长及试管开花诱导
引用本文
张宵娟, 邓光华, 连芳青, 邹娜, 黄颖融, 周树军. 千日红无菌苗生长及试管开花诱导. 草业科学, 2017,34(11):2245-2253
Zhang Xiao-juan, Deng Guang-hua, Lian Fang-qing, Zou Na, Huang Ying-rong, Zhou Shu-jun. Growth and in vitro flowering of Gomphrena globosa . Pratacultural Science,2017,34(11): 2245-2253
Permissions
Zhang Xiao-juan, Deng Guang-hua, Lian Fang-qing, Zou Na, Huang Ying-rong, Zhou Shu-jun. Growth and in vitro flowering of Gomphrena globosa . Pratacultural Science,2017,34(11): 2245-2253
Copyright©2017, 《草业科学》编辑部
千日红无菌苗生长及试管开花诱导
第一作者:张宵娟(1990-),女,江西樟树人,硕士,主要从事园林植物生物技术研究。E-mail:[email protected]
摘要
为进一步探究千日红( Gomphrena globosa)无菌苗生长和试管开花诱导机制,在组培条件下,以带顶芽的千日红茎段为对象,研究培养基中蔗糖含量、外源激素种类(细胞分裂素BA和生长素NAA)及浓度、生长素运输抑制剂(TIBA和NPA)对氮诱导的千日红生长和试管开花的影响。结果表明,蔗糖是千日红无菌苗生长和开花诱导最关键的因素,在不含蔗糖的培养基中,开花率为0且生根较少;随蔗糖含量的增加开花率逐渐提高,并在80 g·L-1时达最大(80.72%);而根系长度和数量及植株长势在蔗糖含量为20~40 g·L-1时较好。BA对无菌苗生长和开花的诱导效应大于NAA,当BA浓度为1.0 mg·L-1时,开花率达最大值60.14%,此时无菌苗长势也较好;而NAA对各指标均无显著性影响。TIBA明显降低了千日红试管开花率并且显著抑制根系的生长,而且在同等条件下对开花率和各生长指标的抑制效应大于NPA。综上所述,氮诱导千日红离体开花和正常生长过程中蔗糖是不可缺少的,并且受外源细胞分裂素的影响和体内激素的调控。
关键词:
千日红; 试管开花; 蔗糖; 细胞分裂素(BA); 生长素(NAA); 生长素极性运输抑制剂
中图分类号:Q943.1
文献标志码:A
文章编号:1001-0629(2017)11-2245-09
doi: 10.11829/j.issn.1001-0629.2016-0642
Growth and in vitro flowering of Gomphrena globosa
Abstract
To clarify the mechanism of growth and in vitro flowering induction of Gomphrena globosa, the shoots with apical bud were used as explants on modified Murashige and Skoog (MS) medium to explore the effects of different concentrations (0, 20, 40, 60, or 80 g·L-1) of sucrose, and different combinations of cytokinins [N6-benzyladenine (BA) and α-naphthaleneacetic acid (NAA)] and auxin polar transport inhibitors TIBA and NPA on nitrogen-regulated growth and flowering in vitro. The results indicated that sucrose was the key factor in floral morphogenesis and growth, and that treatments without sucrose (modified MS supplied with 6.5 g·L-1 agar) produced no flowers and few roots. An increasing concentration of sucrose could promote flowering, and the highest percentage of flowering (80.72%) was obtained on medium containing 80 g·L-1 sucrose. The height, leaf number, and particularly the root volume of G. globosa were all higher in treatments with 20~40 g·L-1 sucrose. L9(34) orthogonal experiment results showed that BA was more effective than NAA in promoting growth and flowering induction, with the highest percentage of flowering (60.14%) and growth being obtained on media containing 1.0 mg·L-1 BA. Auxin polar transport inhibitors prevented flowering and rooting of G. globosa, with the inhibition effects of TIBA being stronger than those of NPA at the same concentrations, although auxin itself did not induce flowering. Therefore, sucrose is indispensable for the growth and in vitro flowering of G. globose. BA also plays an important role, and the effect was also regulated by endogenous hormones.
Keyword:
Gomphrena globosa; flowering in vitro; sucrose; 6-benzyladenine; α; -naphthaleneacetic acid; auxin polar transport inhibitor
千日红为苋科千日红属一年生直立草本植物, 高20~60 cm; 自然条件下6-11月开花, 花色艳丽具光泽, 象征着天长地久、永恒的爱, 因而深受人们喜爱[1]。千日红不仅是天生的干花, 同时又是作鲜切花的良好材料, 而且还可作为花茶饮用。近年来, 从千日红提取天然食用色素和花色苷受到人们普遍关注[2, 3], 诱导千日红开花是其资源开发利用的基础。试管开花由于不受季节限制地周年诱导, 不仅可以达到快繁的目的, 还缩短了开花时间, 可以使新物种和品种更快地进入市场[4]。在前期的研究工作中发现, 接种于改良MS培养基(改变了MS培养基中的N含量及铵硝配比)可以诱导千日红试管苗开花, 并且在培养基中氮总量为5 mmol· L-1, N
1 材料与方法
1.1 试验材料及方法
将千日红无根试管无菌苗接种在生根培养基(1/2 MS+0.5 mg· L-1 IBA+30 g· L-1蔗糖+0.5 g· L-1活性炭), 进行生根壮苗培养[5], 3周后取顶端2 cm(含4片叶)接种到各处理培养基。基本培养基中总氮量为5 mmol· L-1、N
1.1.1 不同蔗糖浓度对千日红无菌苗生长及试管苗开花的影响 采用单因素完全随机试验设计方法, 研究培养基中蔗糖含量(0、20、40、60、80 g· L-1)对千日红无菌苗开花率和生长指标的影响。
1.1.2 不同激素种类及浓度对千日红无菌苗开花诱导和生长的影响 采用L9(34)正交试验设计方法(表1), 研究培养基中不同浓度细胞分裂素BA(0、0.5、1.0 mg· L-1)和生长素NAA(0、0.1、0.5 mg· L-1)对千日红无菌苗开花率和生长指标的影响。
| 表1 植物生长调节剂对千日红试管开花和生长影响的L9(34)正交试验组合 Table 1 Treatment combinations and experimental design for examining the effects of plant growth regulators on the flowering and growth of Gomphrena globosa |
1.1.3 生长素运输抑制剂对千日红无菌苗开花诱导和生长影响 采用单因素完全随机试验设计方法, 研究不同种类和浓度的生长素极性运输抑制剂NPA(1、10 μ mol· L-1)、TIBA(1、10 μ mol· L-1)对千日红无菌苗开花率和生长指标的影响。
1.2 培养条件
培养室温度(25± 1) ℃、每天光照时间12 h, 光照强度为2 000 lx。如无特殊说明各培养基均添加7 mg· L-1亚精胺、琼脂6.5 g· L-1, 蔗糖40 g· L-1, pH 5.8, 121 ℃高压灭菌20 min。
1.3 数据统计与分析
在接种之后的第60天进行苗高和叶片数的测量并统计开花率。苗高用直尺测量, 叶片数为肉眼可见叶片总数, 开花率=开花无菌苗数/接种总数× 100%。每处理分别接种20~30个外植体, 3次重复。应用Excel 2007进行数据处理, 数据的统计分析采用DPS 7.05, 以P< 0.05为差异显著; 采用Duncan新复极差法进行多重比较。
2 结果与分析
2.1 蔗糖对千日红无菌苗开花及生长的影响
观察发现, 无糖培养基中的千日红无菌苗叶片发黄, 基部叶片全部枯黄, 长势较差, 生根少; 随着蔗糖含量的增加, 无菌苗长势逐渐变好, 并且在含有20和40 g· L-1蔗糖的处理中, 千日红无菌苗叶片面积较大且颜色绿, 长势好, 生根数多、根系较长且颜色白; 而在60和80 g· L-1蔗糖处理中的无菌苗基部叶片尖端部分呈黄色, 长势较弱, 无菌苗生根数量逐渐减少, 并且部分根呈黄色, 在80 g· L-1蔗糖处理中的无菌苗根短而粗, 呈黄褐色(图1)。
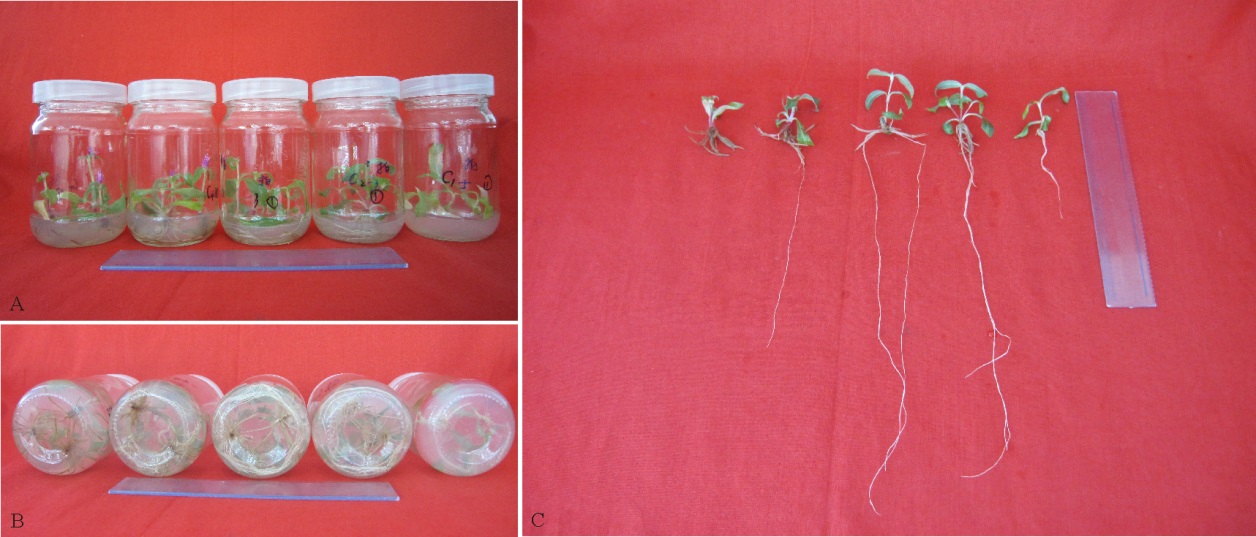 | 图1 蔗糖浓度对千日红无菌苗生长的影响 注:A、B、C表示蔗糖对千日红无菌苗生长(A)和生根(B)及根系生长(C)的影响, 各图中从右到左各植株蔗糖处理浓度分别为0、20、40、60和80 g· L-1。直尺总长为20 cm。Fig. 1 Effects of sucrose on the growth of Gomphrena globosaNote: A, B and C indicate effects of sucrose on growth (A), rooting (B) and root growth (C) of G. globosa. Seedlings in each figures from right to left treated on medium supplied with sucrose of 0, 20, 40, 60 and 80 g· L-1 respectively. Bar=20 cm. |
在接种之后的第22天, 蔗糖含量为60和80 g· L-1的处理中有少部分千日红无菌苗开花, 第45天蔗糖含量为40 g· L-1的处理中也有少部分开花。千日红试管开花率随蔗糖含量的增加呈逐渐上升的趋势(表2), 且在蔗糖含量为80 g· L-1的处理中达到最大值80.72%, 显著高于其他处理(P< 0.05); 而蔗糖对试管苗株高影响差异不显著(P> 0.05); 培养基中添加一定量的蔗糖可以促进试管苗叶片数生长, 并且显著高于对照, 但在含蔗糖各处理间差异不显著。相关性分析结果表明, 开花率与株高和叶片数间的相关系数分别为-0.1和0.353(P> 0.05)。
| 表2 蔗糖含量对千日红试管开花率及生长指标的影响 Table 2 Effects of sucrose content on flowering percentage and growth indices of Gomphrena globosa |
注:同列不同小写字母表示处理间差异显著(P< 0.05)。下同。
Note: Different lowercase letters within the same column indicate significantly different among different sucrose contents at the 0.05 level; similarly for the following tables.
2.2 激素对千日红无菌苗生长及开花诱导的影响
观察发现, 40 d之后, 有部分无菌苗已开花, 60 d时对各处理中开花率、叶片数及株高进行统计(表3)和极差分析(表4)。激素种类对各指标影响的重要性依次为BA> NAA> BA× NAA, 并且在BA为1.0 mg· L-1时千日红无菌苗生长各指标均值最大, 开花率(
| 表3 植物生长调节剂对千日红试管开花和生长影响的正交试验结果 Table 3 Effects of plant growth regulators on the flowering and growth of Gomphrena globosa |
| 表4 植物生长调节剂对千日红试管开花和生长影响的极差分析 Table 4 Range analysis of the effects of plant growth regulators on the flowering and growth of Gomphrena globosa |
注:A, B, C分别为开花率、叶片数和株高; K为各因素水平平均值, R为各因素水平极差、R'为调整后的极差。
Note: A, B and C indicate flowering percentage, leaf numbers and seedling height, respectively; R is range between each factor level; R’ is adjusted range.
方差分析结果表明(表5), BA对千日红试管苗开花率和叶片数的影响达到极显著水平(P< 0.01), 对株高的影响达到显著水平(P< 0.05), 而NAA及二者的交互作用对开花率、叶片数和株高的影响都未达到显著水平(P> 0.05)。植物生长调节剂对千日红试管开花和生长影响的相关性分析结果表明, 开花率与株高和叶片数间的相关系数分别为0.756(P< 0.05)和0.865(P< 0.01)。
| 表5 植物生长调节剂对千日红试管开花率和生长影响的方差分析表 Table 5 Variance analysis of the effects of plant growth regulators on the flowering and growth of Gomphrena globosa |
注:* 表示在0.05水平上差异显著, * * 表示在0.01水平上差异显著。
Note: * and * * indicate significant difference at 0.05 and 0.01 level, respectively.
2.3 生长素极性运输抑制剂对千日红无菌苗开花诱导的影响
处理②和处理③(即添加NPA)的千日红无菌苗开花率仅为对照的44.46%和34.76%(表6), 处理④和处理⑤(即添加TIBA)的开花率仅为对照的29.42%和23.00%。方差分析结果表明, 处理②、③、④、⑤均显著低于对照(P< 0.05), 但处理间差异不显著(P> 0.05)。生长素极性运输抑制剂对千日红试管开花和生长影响的相关性分析结果表明, 开花率与株高和叶片数间的相关系数分别为0.030和0.252(P> 0.05)。
| 表6 生长素极性运输抑制剂对千日红试管开花和生长的影响 Table 6 Effects of auxin polar transport inhibitors on the flowering and growth of Gomphrena globosa |
不同种类及浓度的生长素极性运输抑制剂对千日红无菌苗株高没有显著影响(P> 0.05); 随着NPA处理浓度的增加, 无菌苗生根数逐渐减少、变粗、变短; TIBA处理显著影响无菌苗叶片数和根系生长, 表现为生根数少、根黄色、粗短(图2)。
 | 图2 生长素极性运输抑制剂对千日红无菌苗生长的影响 注:A和B表示生长素极性运输抑制剂对千日红无菌苗生长(A)和生根(B)的影响, 各图中从左到右各处理分别为CK、1和10 μ mol· L-1 NPA、1和 10 μ mol· L-1 TIBA, 直尺总长为20 cm。Fig. 2 Effects of auxin polar transport inhibitors on the growth of G. globose.Note: A and B indicate effects of auxin polar transport inhibitors on growth(A) and rooting(B) of G. globosa. Seedlings in each figures from left to right treated by 0, 1 and 10 μ mol· L-1 NPA, 1 and 10 μ mol· L-1 TIBA respectively. Bar=20 cm. |
3 讨论
3.1 蔗糖对千日红无菌苗生长及开花诱导的影响
糖类作为培养基主要成分之一, 调节植物形态的发生和生理过程[22, 23, 24]。本研究中, 无糖培养基中的千日红无菌苗叶片数量少(表2), 无菌苗生根率低、根系不发达且根的长度较短(图1), 说明蔗糖对千日红无菌苗正常生长具有重要作用[25]。一般在外源蔗糖添加后, 植物体积增加, 可能是通过促进顶端分生组织细胞分裂、增加细胞数量而实现[26]。另外, 蔗糖还可以通过调控地上和地下不同类型的分生组织细胞的活性来影响整个植物的形态, 改变叶片的形状及促进根系的生长[24]。但随着外源蔗糖浓度的增加, 千日红根系生长又受到抑制(图1), 这有可能与高C/N抑制根的生长有关, 并且通过高亲和N
在无糖培养基中, 氮不能诱导千日红无菌苗开花(表2)。根据已有文献报道, 在无糖培养基中, 属间杂种Doriella[10]、杂种茶香月季‘ First Prize’ [14]、白苏[15] 、荞麦(Fagopyrume sculentum)[28]等多种植物的试管开花诱导率为0, 说明蔗糖是多种植物离体开花诱导最关键的因素, 但其启动开花诱导的机制尚不清楚。在三角叶西番莲(Passiflora suberosa)、九里香(Murraya paniculata)、山橘(Fortunella hindsii)等植物上发现蔗糖和光照相互作用可以促进试管开花, Seo等[29]研究发现糖信号很可能是通过下调CONSTANS基因、上调FLOWERING LOCUS T基因的表达, 而成为光周期诱导开花途径的一部分。随着培养基中蔗糖浓度的增加, 千日红试管开花率不断提高, 开花诱导所需要的时间也更短, 这与前人在荞麦[28]、金诺橘(Citrus ‘ Tenora’ ) [30]、吊灯花属(Ceropegia)[31]等多种植物离体开花诱导上的研究结果相一致, 说明高浓度的蔗糖对植物试管开花诱导具有一定的普遍性。这一方面可能是由于C/N对植物开花诱导的影响:在培养基中氮总量保持不变的情况下提高蔗糖含量, 提高了植物的C/N, 促进植株生殖生长, 而在氮供应较多时, 促进蛋白质的合成和植物的营养生长, 但延迟生殖器官的形成[32, 33]。另一方面, 增加培养基中蔗糖含量, 也可能是通过其渗透作用调节千日红无菌苗开花诱导, 有研究表明, 高浓度的凝固剂会提高带节荞麦茎段的离体开花诱导率[28]。但Taylor等[34]的研究结果表明, 培养基中碳水化合物对火炬花(Kniphofia leucocephala)离体开花诱导效应似乎直接来自蔗糖而不是其渗透作用, 因为相应的甘露醇并不能诱导开花; 碳水化合物通过其渗透效应促进开花的可能性, 也被在白芥菜(Sinapsis alba)的研究中所否定[35]。另外, 与在火炬花上的研究相似[34], 千日红试管开花率与生长各参数之间缺乏明显的相关性(P> 0.05), 表明蔗糖对开花诱导的促进并不是来自对植物整体生长的刺激, 而是对离体开花诱导的直接效应。
3.2 植物生长调节剂对千日红无菌苗生长和开花诱导的影响
细胞分裂素因为调节细胞的分裂和器官的形成而被认为是植物离体开花所必需[36]。本研究结果表明, 细胞分裂素BA对千日红试管开花诱导有极显著性影响, 并且在BA为1.0 mg· L-1时开花率达最大值60.14%。外源细胞分裂素对离体开花诱导的促进效应也已在荞麦[28]、吊灯花属[31]和火炬花[37]等多种植物中广泛报道。研究证实, 在花芽分化后, 苗顶端分生组织中细胞分裂素供应量或含量增加[37, 38]。根尖是细胞分裂素合成的主要部位, 其经木质部运输至地上器官, 通常认为由根到茎的细胞分裂素信号调控植物的营养生长向生殖生长的转变[39]。一些研究表明, 在硝铵共培养条件下植物体内细胞分裂素含量较高[40], 添加N
3.3 生长素极性运输抑制剂对千日红无菌苗生长开花诱导的影响
本研究中虽然NAA本身并不诱导开花, 经生长素运输抑制剂处理后, 千日红无菌苗开花率显著下降, 这一现象与Kostenyuk等[11]对兰花的研究结果一致, 生长素运输抑制剂降低植物试管开花率的机制目前尚不清楚, 可能与生长素能够调控细胞分裂素在植物某些组织中的合成有关[43]。此外, 经NPA和TIBA处理的千日红无菌苗根系生长也受到显著抑制, 类似的现象在挪威云杉(Picea abies)离体幼苗上也有报道[44]。生长素主要在植物体中具有分生能力的茎尖、幼叶和根等组织器官中合成, 向根尖极性运输, 并由极性运输所形成的生长素浓度梯度参与调控植物的许多生理过程, 成为植株形态的决定者[45]。分子遗传学和生理学研究表明, 生长素极性运输取决于在细胞极性分布的输入和输出载体, 而生长素运输抑制剂通过抑制IAA与载体蛋白的结合[46], 从而抑制生长素的极性运输。本研究结果表明, 植株体内生长素极性运输在千日红试管花诱导和正常发育过程中起重要作用, 氮可能通过对激素信号、激素运输或激素合成的调控, 进而影响植物的生长和开花诱导, 如生长素受体TIR3和一些生长素载体蛋白的转录水平受氮调控[47]。
4 结论
本研究结果表明, 千日红离体开花和正常生长过程中蔗糖是不可缺少的; 并且在蔗糖含量为80 g· L-1时开花率达最大, 80.72%, 而植株整体长势在蔗糖含量为20~40 g· L-1时较好, 表明蔗糖对开花诱导的促进并不是来自对植物整体生长的刺激, 而是对离体开花诱导的直接效应, 并且与C/N有关。BA在千日红试管开花和生长过程中起重要作用, 且浓度为1.0 mg· L-1时, 无菌苗开花率达最大值, 60.14%, 此时无菌苗整体生长势也较好; 经生长素运输抑制剂处理后, 千日红无菌苗开花率显著下降、根系生长受抑制, 表明千日红试管开花和生长发育过程受体内外激素影响。因此, 氮对千日红生长和试管开花诱导的影响与C/N比和激素调控有关。
The authors have declared that no competing interests exist.
参考文献
| [1] |
|
| [2] |
|
| [3] |
|
| [4] |
|
| [5] |
|
| [6] |
|
| [7] |
|
| [8] |
|
| [9] |
|
| [10] |
|
| [11] |
|
| [12] |
|
| [13] |
|
| [14] |
|
| [15] |
|
| [16] |
|
| [17] |
|
| [18] |
|
| [19] |
|
| [20] |
|
| [21] |
|
| [22] |
|
| [23] |
|
| [24] |
|
| [25] |
|
| [26] |
|
| [27] |
|
| [28] |
|
| [29] |
|
| [30] |
|
| [31] |
|
| [32] |
|
| [33] |
|
| [34] |
|
| [35] |
|
| [36] |
|
| [37] |
|
| [38] |
|
| [39] |
|
| [40] |
|
| [41] |
|
| [42] |
|
| [43] |
|
| [44] |
|
| [45] |
|
| [46] |
|
| [47] |
|