

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
干旱-复水对菊芋苗期根、茎、叶形态特征的影响
引用本文
朱铁霞, 王琳, 高阳, 王伟东, 高凯. 干旱-复水对菊芋苗期根、茎、叶形态特征的影响. 草业科学, 2017,34(11):2309-2315
Zhu Tie-xia, Wang Lin, Gao Yang, Wang Wei-dong, Gao Kai. Effect of drought stress and re-watering on the morphological characteristics of root, stem, and leaf in Helianthus tuberosus . Pratacultural Science,2017,34(11): 2309-2315
Permissions
Zhu Tie-xia, Wang Lin, Gao Yang, Wang Wei-dong, Gao Kai. Effect of drought stress and re-watering on the morphological characteristics of root, stem, and leaf in Helianthus tuberosus . Pratacultural Science,2017,34(11): 2309-2315
Copyright©2017, 《草业科学》编辑部
干旱-复水对菊芋苗期根、茎、叶形态特征的影响
第一作者:朱铁霞(1978-),女,吉林通化人,讲师,硕士,主要从事草地资源与利用研究。E-mail:[email protected]
摘要
通过盆栽试验研究干旱-复水对菊芋( Helianthus tuberosus)苗期根、茎、叶、茎叶比、根冠比和物质积累量的影响,探讨菊芋苗期干旱适应机制,干旱半干旱地区菊芋栽培管理提供参考。结果表明:干旱-复水处理菊芋根、茎、叶生物量和总生物量均低于正常水分管理;干旱-复水增加了主根和须根的直径;干旱-复水过程中除复水第16天外,处理须根长度均高于对照;处理前期主根长度高于对照,后期主根长度低于对照;干旱-复水处理提高了菊芋平均叶面积,降低了叶片数量,抑制新叶片的形成;干旱胁迫第2天测定的茎叶比对照高于处理,而其他取样时间处理均高于对照;干旱-复水过程中菊芋根冠比胁迫第2天高于对照,胁迫第4天、第8天和复水第8天处理根冠比低于对照,复水第16天处理高于对照。
关键词:
菊芋; 干旱-复水; 根; 茎; 叶
中图分类号:Q943.1
文献标志码:A
文章编号:1001-0629(2017)11-2309-07
doi: 10.11829/j.issn.1001-0629.2017-0007
Effect of drought stress and re-watering on the morphological characteristics of root, stem, and leaf in Helianthus tuberosus
Abstract
Using pot experiments, we examined the effects of drought stress and re-watering on root, stem, leaf, stem/leaf ratio, root/shoot ratio, and biomass in Helianthus tuberosus The following results were obtained. Root, stem, leaf biomass, and total biomass in treated plants were lower than those in control plants. The diameter of the taproot and fibrous roots was increased. During drought stress and re-watering, the lengths of fibrous roots were greater than those in control plants, and the length of the taproot was greater than the control during drought stress but lower than the control during re-watering. The mean leaf area of treated plants was higher than that of control plants, and the number of leaves were lower than that in control plants. The stem/leaf ratios of treated plants were all higher than those of control plants, except on the second day of measurement. During drought stress and re-watering, the root/shoot ratio of treated plants under drought stress on the second day of measurement and re-watered plants at 16 days was higher than that of the control plants, but lower than that of control plants on the fourth and eighth days under drought stress and eighth day under re-watering.
Keyword:
Helianthus tuberosus; drought stress and re-watering; root; stem; leaf
植物根、茎、叶等器官的形态特征与生长环境条件密切相关, 在适应外界环境过程中, 各器官形态结构表现出一定的变异性和可塑性, 这种变化对外界环境的反应和适应最早也最为明显[1]。而在作物栽培过程中, 长期干旱的情况极少, 往往干旱-复水交互出现的情况较多。因此, 研究干旱-复水对作物的影响意义更大。
菊芋(Helianthus tuberosus)又名洋姜、鬼子姜, 菊科向日葵属, 多年生草本植物。原产北美, 经欧洲传入中国, 其块茎菊粉含量丰富, 约占干物质70%~90%[2]。菊粉是果糖的聚合物, 外切型菊粉酶将其分解为单个果糖, 再经发酵生产乙醇, 转化率高达83%~99%[3, 4, 5], 并且其对生境要求低, 具有喜肥耐贫瘠, 喜湿耐旱, 喜温耐寒, 耐盐碱等优点, 在自然环境较差, 无法进行农作物生产的区域具有一定栽培优势, 并且菊芋具有种植成本低廉、生物产量高的优点。因此, 菊芋将是燃料乙醇和菊粉产业规模化发展可选择的重要植物, 其生产战略意义重大, 应用前景广阔。近年来, 我国菊芋种植面积呈现逐年增加的趋势, 尤其在干旱和半干旱地区。然而, 菊芋的研究主要集中于菊芋的收获时间、肥料管理、种植密度、水分管理等相关内容[6, 7, 8, 9, 10, 11, 12], 关于菊芋干旱适应机制的研究却很少, 只有少数学者针对干旱胁迫条件下菊芋苗期叶片保护酶活性、生长节律和光合特性等相关内容进行了初步研究[13, 14]。截至目前, 对菊芋干旱适应机制进行系统的、全面的阐述还鲜见报道。然而菊芋干旱适应机制的研究, 是其在干旱地区种植、推广的前提条件, 是旱区植物栽培、管理的重要依据, 其研究意义重大。基于上述原因, 本研究拟通过对干旱-复水条件下根、茎、叶形态特征的研究, 探讨菊芋抗旱生理机制, 为菊芋旱地栽培管理提供依据。
1 材料与方法
1.1 试验设计
试验于2016年5月-2016年8月在内蒙古民族大学农学院试验农场(43° 36' N, 122° 22' E)遮雨棚下进行。采用盆栽方式种植, 花盆直径29 cm, 高度32 cm。装盆土壤为灰色草甸土, 是当地主要土壤类型, 装盆前将土壤混合均匀, 每盆装入土壤重量相同。土壤有机质含量为18.23 g· kg-1、碱解氮含量为62.41 mg· kg-1、速效磷含量为38.61 mg· kg-1、速效钾含量为184.56 mg· kg-1、pH 8.2。菊芋品种为红皮菊芋, 种植时选取重量为30~40 g, 无病、无伤的块茎作种。
干旱胁迫处理从30苗龄(营养生长期, 株高20~25 cm, 叶片数量6~8枚。)开始, 1-30苗龄之间保证菊芋正常生长, 没有受到干旱或者其他逆境胁迫。干旱处理分为两个阶段, 第一阶段为持续干旱胁迫阶段, 即不对菊芋进行灌水, 第二阶段为复水阶段, 即持续干旱胁迫8 d后对菊芋进行补水, 复水后每天确保菊芋生长的正常需水量, 每天约补水1 L, 避免菊芋受到两次干旱胁迫。其中干旱处理20盆, 以正常灌水为对照, 20盆。
样品采集时间分别为干旱胁迫后第2天、第4天、第8天和复水后第8天和第16天, 每次取处理和对照各4盆, 共计8盆。将菊芋植株齐根割取地上部分, 进行茎、叶分离。同时, 将地下根系进行清洗、取样。
1.2 测定指标与方法
生物量:每次取样将整株齐根刈割, 地上部分进行茎、叶分离, 地下部分通过清洗去土, 尽量取出完整根系。将所取样品在105 ℃下杀青30 min, 在(75± 5) ℃条件下烘干, 称取干重。
叶片数量记录:取样前对整株叶片进行计数。
单片叶面积:利用AM350便携式叶面积仪对全株叶片进行叶面积测定, 除以全株叶片总数, 得出平均单片叶叶面积。
根冠比=地下(根)生物量/地上生物量;
茎叶比=茎干重/叶片干重。
1.3 数据分析
利用Excel完成数据计算, 利用SPSS 17.0对同时期取样的对照和处理进行单因素方差分析。
2 结果与分析
2.1 干旱-复水对生物量的影响
胁迫第2天和复水第8天对照叶干重显著高于处理(P< 0.05), 胁迫第2天、复水第8天和复水第16天对照茎重量显著高于处理, 胁迫第8天和复水第16天对照根干重显著高于处理, 复水第8天处理根重量显著高于对照, 胁迫第2天、复水第8天和复水第16天处理总生物量显著低于对照(P< 0.05)(表1)。
| 表1 干旱-复水对红皮菊芋苗期生物量的影响 Table 1 Effect of drought stress and re-watering recovery on seedling biomass of Jerusalem artichoke |
注:同列不同小写字母表示同一指标处理与对照间差异显著(P< 0.05)。
Note: Different lowercase letters with in the same column indicate significant difference between control and drought stress treatment within the same parameter at the 0.05 level.
2.2 干旱-复水对叶面积的影响
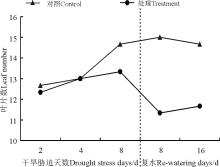
菊芋叶面积除胁迫第8天和复水第16天对照和处理之间没有显著差异外(P> 0.05)(图1), 其他处理单叶叶面积均显著高于对照(P< 0.05); 红皮菊芋单叶叶面积伴随干旱-复水呈现“ 降-升-降” 的变化趋势, 对照叶面积随时间延长呈现“ W” 型变化。
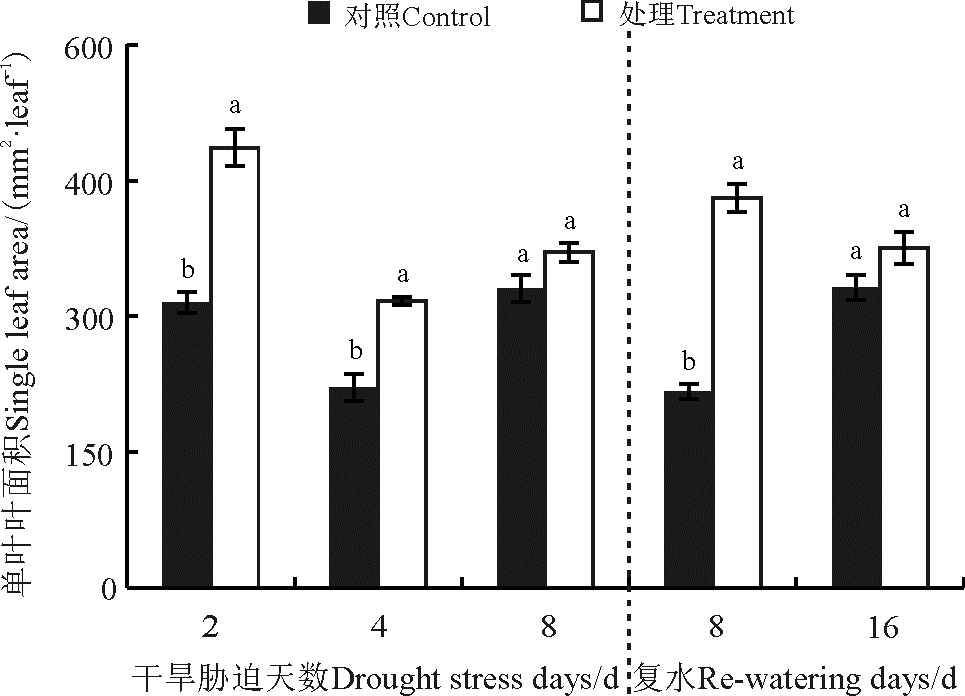 | 图1 干旱-复水对菊芋单叶叶面积的影响 注:不同小写字母表示差异显著(P< 0.05)。下同。Fig. 1 Effect of dorught and re-watering on leaf area in Hellanthus tuberosusNote: Different lowercase letters indicated significantly difference between control and treatments at the 0.05 level; similarly for the following figures. |
2.3 干旱-复水对叶片数的影响
对照叶片数量从胁迫第8天到复水第16天均高于处理, 对照叶片数前期逐渐增加, 后期变化比较平缓, 处理叶片数量从胁迫第2天到胁迫第8天变化比较平缓, 胁迫第8天到复水第8天叶片数量大幅度下降, 复水第8天到复水第16天呈现上升趋势(图2)。
 | 图2 干旱-复水对菊芋叶片数量的影响Fig. 2 Effect of dorught and re-watering on leaf number in Hellanthus tuberosus |
2.4 干旱-复水对根系长度和直径的影响
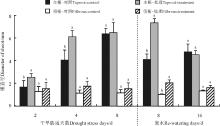
菊芋主根长度在胁迫第2天、胁迫第4天和复水第8天处理高于对照(图3), 且胁迫第4天和复水第8天显著高于对照(P< 0.05), 胁迫第8天和复水第16天对照高于处理, 且复水第16天显著高于对照; 复水第16天须根长度对照高于处理(P> 0.05), 其他处理均高于对照, 其中胁迫第8天和复水第8天处理显著高于对照。
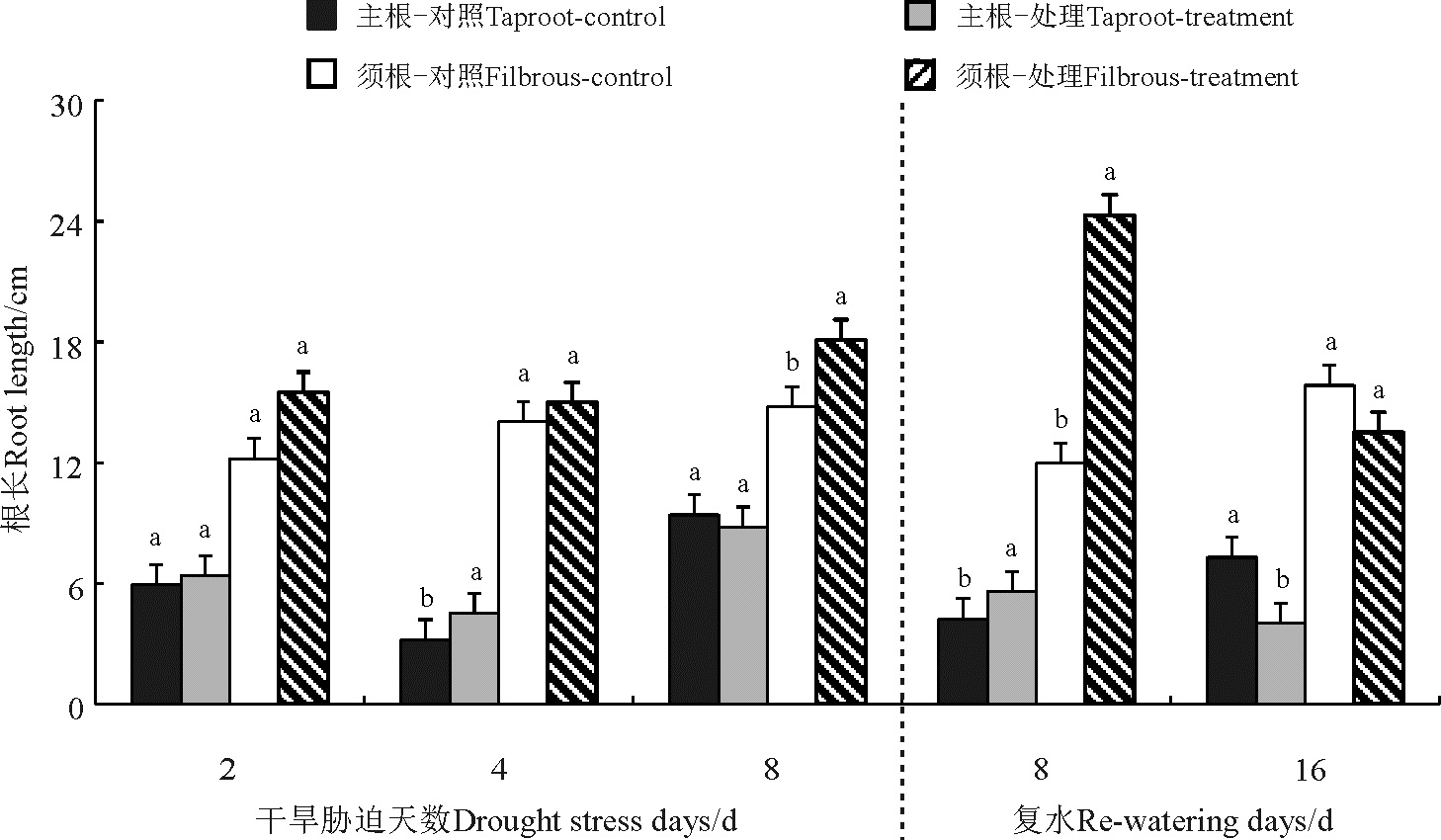 | 图3 干旱-复水对菊芋苗期主根和须根长度的影响Fig. 3 Effect of dorught and re-watering on taproot and fibrous lenght of Hellanthus tuberosus |
主根直径除了复水第16天对照高于处理之外(P> 0.05), 其他均为处理高于对照(图4), 且胁迫第2天、第4天和复水第8天处理显著高于对照(P< 0.05); 须根直径处理均高于对照, 其中复水第8天显著高于对照。
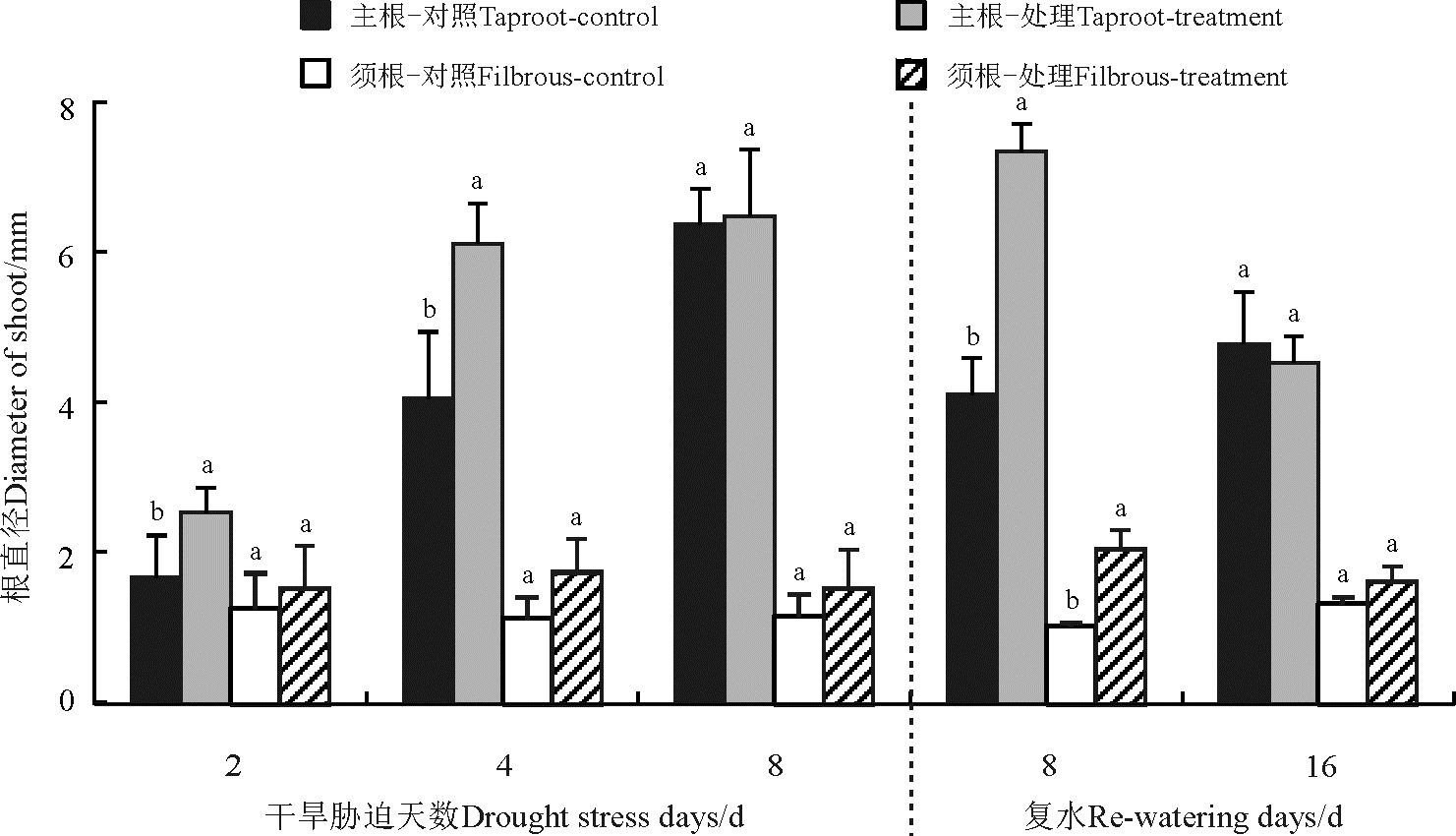 | 图4 干旱-复水对菊芋苗期主根和须根直径的影响Fig. 4 Effect of dorught and re-watering on taproot and fibrous diameter of Hellanthus tuberosus |
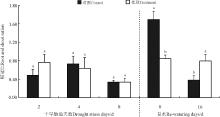
2.5 干旱-复水对茎叶比的影响
干旱-复水处理后菊芋茎叶比除了胁迫第2天对照高于处理之外, 其他各处理均高于对照(图5), 其中胁迫第4 天、复水第8天和复水第16天显著高于对照(P< 0.05), 随胁迫时间的延长, 处理茎叶比呈现“ 升-降-升-降” 的变化趋势, 峰值分别出现在胁迫第4天和复水第8天。
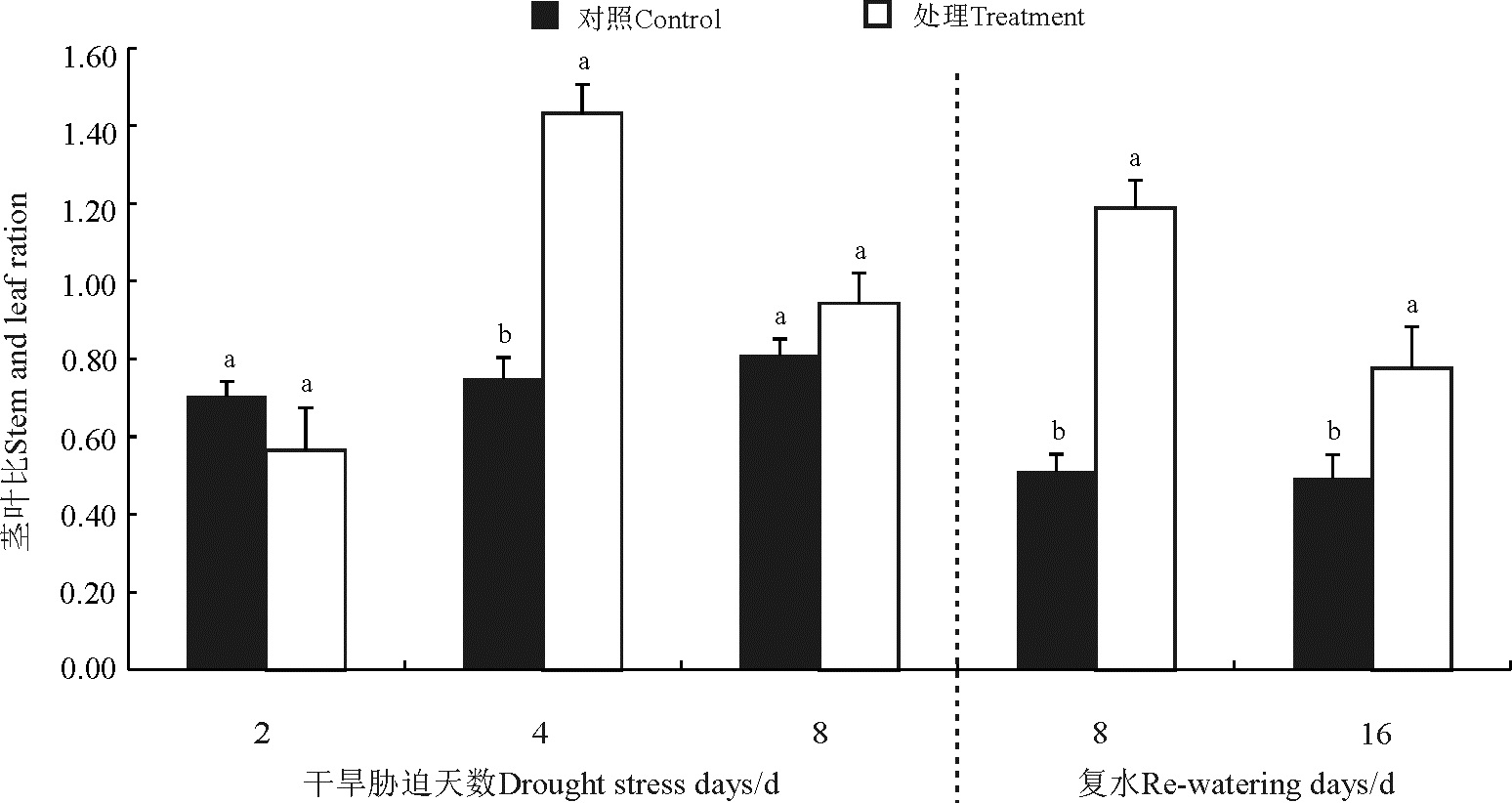 | 图5 干旱-复水对菊芋茎叶比的影响Fig. 5 Effect of dorught and re-watering on stem/leaf ratio of Hellanthus tuberosus |
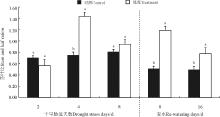
2.6 干旱-复水对根冠比的影响
胁迫第4天、第8天和复水第8天根冠比对照高于处理, 其中复水第8天根冠比对照显著高于处理(P< 0.05), 胁迫第4天和第8天处理和对照间无显著差异(P> 0.05), 胁迫第2天和复水16天处理高于对照, 其中复水第16天处理显著高于对照(P< 0.05), 胁迫第2天对照和处理差异不显著(图6)。
 | 图6 干旱-复水对菊芋根冠比的影响Fig. 6 Effect of dorught and re-watering on root/shoot ratio of Hellanthus tuberosus |
3 讨论
干旱-复水试验作为作物干旱机制研究的热点内容, 已在玉米(Zea mays)、大豆(Glycine max)、小麦(Triticum aetivum)等作物的进行了研究, 研究证实了干旱阈值内进行干旱胁迫并及时复水, 能够促使作物的株高、叶面积和生物量等超过正常灌水水平, 即“ 补偿或超补偿效应” [15]。然而, 在干旱-复水过程中植株的茎、叶、根等器官的形态会表现出一定的变化。如, 水稻(Oryza sative)在干旱-复水的研究过程中, 发现经过处理能够诱导水稻根系变长, 且胁迫越重根系越长, 同时, 根系的重量增加[16]。本研究中对30苗龄菊芋进行干旱和复水处理, 通过主根和须根长度、直径的测定发现, 须根对干旱-复水的响应更为敏感, 其中除复水第16天外, 其他处理须根长度均高于对照; 主根长度胁迫处理前期高于对照, 后期低于对照; 干旱-复水对主根和须根直径的影响比较一致, 除复水第16天主根直径外, 其余处理均表现为处理高于对照。根系变长变粗是对干旱胁迫的一种适应性表现, 根系变长是为了增加吸水范围, 以便根系能够吸收地下更深层的水来降低干旱胁迫的影响, 变粗能够有效地提高自身的储水能力, 也是降低干旱胁迫的一种形态学适应, 须根比主根对干旱-复水处理更为敏感, 可能主要有两个原因, 第一, 须根相对主根属于新生根, 其对环境的反应比较敏感; 第二, 须根是植物吸收水分和营养物质的主要部位, 因此, 其对外界环境的响应更为积极。
叶片是植物光合作用的主要器官, 其形态与外界环境关系密切。本研究结果表明, 干旱-复水处理提高了菊芋平均单片叶叶面积, 降低了叶片数量。主要是因为干旱-复水处理抑制了新生叶片的形成, 促使叶片平均叶面积高于对照。该结论也说明植物苗期受到干旱胁迫会通过减少新生叶片数量, 来降低叶片的蒸腾耗水, 从而提高植株对外界干旱环境的耐受能力。同时, 通过叶片数量变化可以看出, 从复水第8 天到复水第16 天, 叶片的数量开始增加, 但增加幅度比较平缓, 这表明植物干旱胁迫后及时复水植株能够表现出一定的补偿生长迹象。同时, 通过茎叶比可以看出, 干旱胁迫第2 天测定的茎叶比对照高于处理, 而其他取样时间处理均高于对照, 主要是由于干旱-复水胁迫抑制了新生叶片的形成, 从而降低了叶片的生物量, 导致茎叶比高于对照。
根冠比是指植物地下部分与地上部分的鲜重或干重的比值。它的大小反映了植物地下部分与地上部分的相关性, 外界生长环境条件对其有一定的影响[17]。本研究在干旱-复水过程中菊芋幼苗根冠比胁迫第2天初期处理高于对照, 胁迫第4 天、第8 天和复水第8 天处理低于对照, 复水第16 天处理高于对照。该结论与郝树荣等[16]对水稻干旱-复水试验的研究结论相似, 均为干旱胁迫降低根冠比, 且胁迫越重, 根冠比越小, 复水后根冠比呈现增加的趋势; 与其他学者[18, 19, 20]对苗木水分胁迫后根冠比的变化略有差异, 该研究认为干旱胁迫促使苗木根系生物量增加, 导致根冠比增加, 有助于苗木吸收更多的水分。导致研究结论不同的原因可能为, 研究对象的不同、植物生育期不同等。
The authors have declared that no competing interests exist.
参考文献
| [1] |
|
| [2] |
|
| [3] |
|
| [4] |
|
| [5] |
|
| [6] |
|
| [7] |
|
| [8] |
|
| [9] |
|
| [10] |
|
| [11] |
|
| [12] |
|
| [13] |
|
| [14] |
|
| [15] |
|
| [16] |
|
| [17] |
|
| [18] |
|
| [19] |
|
| [20] |
|