

{kind=link}
{kind=link}
{kind=link}
灌溉条件下不同紫花苜蓿品种生长动态模型及其类型
引用本文
祁翠兰, 武得礼. 灌溉条件下不同紫花苜蓿品种生长动态模型及其类型. 草业科学, 2017,34(3):556-564
Qi Cui-lan, Wu De-li. Growth model and classification of different alfalfa varieties under irrigation conditions. Pratacultural Science,2017,34(3): 556-564
Permissions
Qi Cui-lan, Wu De-li. Growth model and classification of different alfalfa varieties under irrigation conditions. Pratacultural Science,2017,34(3): 556-564
Copyright©2017, 《草业科学》编辑部
《草业科学》编辑部
灌溉条件下不同紫花苜蓿品种生长动态模型及其类型
摘要
研究了灌溉条件下15个紫花苜蓿( Medicago sativa)品种二年龄第一茬草的生长动态,其动态模型符合Logistic生长曲线;各品种之间的回归模型有明显差别。依生长曲线方程计算了各品种的最高产量潜力(K),最大生长速度(Vmax)、生长最快时间(t)、生长速度由慢变快的突变点(t1)和由快变慢的突变点(t2)、旺盛生长期(t2-t1)和生长特征值(GT)。t2可作为刈割始期的指标,供生产中参考应用。对这些特征值进行了偏相关分析,表明苜蓿产量潜力的大小取决于旺盛生长期和最大生长速度二者的乘积。以这些特征值为依据进行了聚类分析,第二类品种(金皇后、巨人201、德宝、三得利、维拉和瑞西)表现良好,宜在当地推广种植,刈割始期为5月17日。
关键词:
紫花苜蓿; 生长曲线; 聚类分析; 刈割始期
中图分类号:S816
文献标志码:A
文章编号:1001-0629(2017)3-0556-09
doi: 10.11829/j.issn.1001-0629.2016-0176
Growth model and classification of different alfalfa varieties under irrigation conditions
Abstract
The growth model for 15 varieties of alfalfa as the first crop in the second year under irrigation conditions was investigated. The results showed that the growth model was consistent with the logistic growth curve. There were obvious differences in the regression models of varieties. Based on these models, the highest potential yield (K), maximum growth speed (Vmax), time of maximum growth (t), turning point of growth speed from slow to fast, and that of fast to slow (t1, t2), the period of vigorous growth (t2-t1), and growth characteristic values (GT) were calculated. The variable t2 can be used as an index to the time to begin harvest with reference to production. Partial correlation analysis showed that the potential yield of alfalfa was determined by the product of days of vigorous growth and maximum growth speed. According to the values, the varieties were clustered into five types. The second classification varieties (Golden Empress, Ameristand 201, Derby, Sandili, Vela, and Rest) are appropriate for local planting with harvest beginning on May 17.
Keyword:
alfalfa; growth curve; cluster analysis; beginning time for harvest
紫花苜蓿(Medicago sativa)是世界著名的优良多年生豆科牧草, 在畜牧业生产中发挥着非常重要的作用。长期以来, 很多科研工作者对紫花苜蓿进行了大量的研究, 特别是21世纪以来, 由于国外苜蓿新品种的大量引进, 许多学者进行了苜蓿新品种的适应性、生物量特性及生产性能评价的研究, 如在河西走廊、陇中和陇东黄土高原丘陵区都进行了紫花苜蓿新品种的引种和适应性研究[1, 2, 3], 在辽西北风沙半干旱区、贵州南部和冀西北地区也进行了紫花苜蓿新品种的适应性和生长特性的研究[4, 5, 6, 7, 8]。由于各地的生态条件差异大, 所适宜的品种也各不相同。景泰高扬程提灌区是一个新灌区, 紫花苜蓿种植的历史尚短, 在品种的选择、栽培管理及刈割利用等方面, 都缺乏系统的研究, 生产存在着极大的盲目性, 不利于苜蓿生产的进一步发展。因此, 引进并选择适宜当地条件的紫花苜蓿新品种, 并进行生长特性及栽培管理方面的系统研究, 对促进当地的畜牧业发展, 具有重要的意义。
苜蓿的地上生物量是人们要获取的目标产物, 生物量特性是苜蓿最重要的生长特性之一。研究苜蓿生物量的动态变化, 并用数学模型进行定量模拟, 通过数学的方法对其研究, 可以准确地描述不同苜蓿品种的差异。关于苜蓿地上生物量积累动态数学模型的研究报道较少。目前的研究内容可归纳为三类:一是研究了紫花苜蓿地上生物量依返青后天数变化的数学模型[9, 10, 11]; 二是研究了紫花苜蓿植株高度随生育天数变化的动态模型[12, 13]; 三是研究了紫花苜蓿地上干物质产量与积温之间的关系[14]。上述各项研究仅进行了生长曲线模型的拟合, 对不同品种生长动态的比较, 并用生长曲线的特征值进行聚类和相关分析的研究未见报道。本研究通过对不同苜蓿品种生长动态的模拟, 进而对生长曲线的特征值进行相关分析和聚类分析, 以揭示各品种的生长规律, 用于指导当地的紫花苜蓿生产。
1 材料和方法
1.1 试验地概况
试验设在兰州大学草地农业科技学院景泰试验站, 位于104° 30' E, 37° 10' N, 海拔1 630 m, 年平均气温8.2 ℃, 无霜期141 d, ≥ 10 ℃活动积温3 038.2 ℃· d, ≥ 0 ℃活动积温3 614.8 ℃· d; 土壤类型为灰钙土, 灌溉条件良好, 代表该地区中上等地力水平。
1.2 供试材料
共种植紫花苜蓿品种15个, 其中国外引进品种11个, 国内品种4个(表1), 所有品种来自兰州大学草地农业科技学院。
| 表1 供试品种 Table 1 Varieties used in the study |
1.3 试验方法
采用完全随机区组设计, 2次重复, 小区面积4 m× 6 m, 2013年6月16日播种, 人工开沟条播, 行距33 cm, 6月21日灌水, 23日出苗, 出苗率90%以上, 以后管理同一般大田。2013年未收割。2014年春季仔细观察返青情况, 从返青期3月28日开始, 每7 d测定一次, 至5月30日开花期结束, 从地上3~5 cm处刈割, 样品装入塑料袋, 在室内立即称鲜重; 取小样本(前3次未取小样本, 样品全部处理)约1 000 g称重后在105 ℃下杀青15 min, 再在80 ℃下烘干至恒重, 称干重并折算产量。取样面积为0.9 m2, 每小区取1个样方。
1.4 数据统计分析方法
用Excel作散点图进行直观对比, 用SAS统计分析软件中Reg过程进行回归方程的计算, 用ANOVA过程进行方差分析, 用Corr过程计算偏相关系数, 用Cluster过程进行聚类分析(数据的标准化采用标准差标准化)。
2 结果与分析
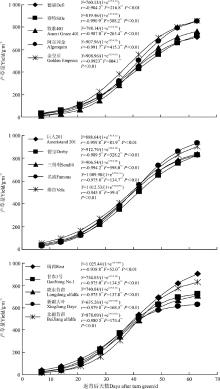
紫花苜蓿的地上生物量随返青后天数的增加而增加(表2), 前期增长较慢, 中期增速加快, 后期增速又变慢, 巨人201、名流、维拉、瑞西、甘农1号、陇东苜蓿、北疆苜蓿7个品种在56 d后生物量出现下降。各品种之间的增速和生物量有一定差异。牧歌401、阿尔冈金在返青初期的干重最小, 之后生长速度加快, 到后期产量较高。维拉、瑞西等品种返青初期就表现产量高, 中后期一直处于高水平。新疆大叶、陇东苜蓿等国内品种, 在返青初期虽然产量不低, 中期的产量水平也与其它品种相当, 但后期生长速度较慢产量较低。
| 表2 产草量(g· m-2)测定结果 Table 2 Results of measurements of grass yield (g· m-2) |
2.1 生长动态模型
对于紫花苜蓿生物量积累过程数学模型的研究报道较少, 相关研究表明, 不同秋眠级苜蓿品种地上总生物量积累的动态模型符合“ S” 型Logistic生长曲线[15]。另有研究表明, 不同苜蓿品种在返青后的生长高度呈直线上升, 没有表现出明显的慢-快-慢的生长节律[16]。在陕西关中地区对5个苜蓿品种地上生物量的绝对生长量(AGR)和相对生长量(RGR)进行研究, 返青初期温度较低, AGR较小; 4月中旬后随水热同期到来, AGR达到最大, 随后因生殖生长的影响, AGR下降, 表现出明显的慢-快-慢的生长规律[17]。在作物生长后期, 由于不良环境条件和生殖生长的影响, 作物为了维持生命, 积累的干物质又转化而被作物消耗, 干物质可能有下降的趋势, 这种情况必须用修正的生长曲线来进行拟合[18]。可见, 在不同的生长条件下, 苜蓿干物质积累的动态也可能不同。因此, 我们用直线方程Y=a+bX、生长曲线Y=K/(1+ea+bX)和修正生长曲线Y=K/(1+
| 表3 三种数学模型的离回归标准误和F测验值 Table 3 Standard error of estimate and F for 3 math. model |
 | 图1 不同苜蓿品种的生长曲线Fig.1 Growth curve of different varieties of alfalfa |
2.2 对生长曲线的数学分析
Logistic曲线是一个“ S” 型曲线。在作物生长初期, 生长速度慢, 之后由慢逐渐变快, 在一定时间生长速度达到最大值, 此时产量水平为K/2, 之后生长速度逐渐变慢, 并逐渐趋于0, 产量水平逐渐趋近于K值。K值的大小, 表明了该品种最大生产潜力的大小。
对生长曲线方程求一阶导数, 可得到生长速度方程:
V=K(-b)ea+bx/(1+ea+bx)2.
它是一个连续变化的单峰曲线。依此方程可求出最大生长速度出现的时间t=-a/b, 此时的最大生长速度为Vmax=-bK/4, 它们分别表示了苜蓿生长最快时出现的时间和生长速度的最大值, 它们都是重要的品种特性。据此方程还可求出生长速度由慢变快的突变点t1和由快变慢的突变点t2; t1出现的早晚, 表示一个苜蓿品种返青后生长的快慢, 与早期产量有直接关系; t2则表示苜蓿的生长由快速生长阶段进入缓慢生长阶段, t2出现的迟早, 表示刈割时期的早晚。t2- t1则表示作物旺盛生长期的长短, 它与Vmax的乘积称为生长特征值(GT), 它表示作物在旺盛生长期的生长量。
根据以上分析, 分别计算了各品种的最大生长速度Vmax、最大速度出现的时间t、生长速度由慢变快的突变点t1和由快变慢的突变点t2、旺盛生长期的长短t2-t1、生长特征值GT。苜蓿的最大生产潜力以瑞西最大, 维拉、名流次之, 新疆大叶最小, 甘农1号和陇东苜蓿也较小(表4)。究其原因, 国内品种虽然t1值较小, 前期发育早, 但t2值也小, 快速生长期结束也早, 旺盛生长期较短, 生长速度也较小, 因而产量潜力较低。
| 表4 生长曲线的特征值 Table 4 Characteristic value of growth curves |
2.3 苜蓿生长的内在规律性分析
根据苜蓿生长的动态模型所求出的各个特征值, 反映了一个苜蓿品种的某些本质属性; 对它们进行深入分析, 可以揭示苜蓿生长的某些内在规律性。将表4中的各个特征值之间进行了偏相关分析(表5), 结果表明:K值与GT有极显著的正相关(P< 0.05), 而与其它各项均无相关; 说明苜蓿生产潜力的大小, 并不单纯决定于最大生长速度的大小, 与最大生长速度出现的迟早及旺盛生长期的长短无关, 而是取决于最大生长速度与旺盛生长期二者的乘积。t1与t之间相关极显著, 说明返青后生长越快的品种, 最大生长速度出现的也越早。t与t2之间有显著正相关(P< 0.05), 表明最大生长速度出现较迟的品种, 生长由快变慢的转折点也相应较迟。但t1与t2之间无相关, 说明2个突变点的变化并不是同步的。t1与Vmax之间有显著正相关, 表明前期生长发育较迟缓的品种, 最大生长速度较大。t2与t2-t1之间也有显著正相关, 表明后期发育较迟的品种, 它的旺盛生长期较长。
| 表5 各特征值之间的偏相关系数 Table 5 Coefficient of partial correlation between characteristic values |
2.4 聚类分析
各个苜蓿品种的生长动态曲线, 有些比较相似, 有些则相差较大。通过聚类分析, 把比较相似的品种进行适当归类, 则更便于分析和生产中应用。以表4中所列的7个特征值为分类指标, 进行了系统聚类。以连接距离0.75进行了划分, 可分为5类(图2):
 | 图2 聚类分析结果注:1-15为品种序号, 同表1。Fig.2 Results of cluster analysisNote:1-15 are codes of varietives, the same in Table 1. |
第Ⅰ 类:包括牧歌401、阿尔冈金、名流和北疆苜蓿, 前期生长慢, t1出现在返青后29 d; 最大生长速度最大, 达26.9 g· (m2· d)-1; 但旺盛生长期较短, 因而产量水平属中上水平。属于前慢后快的类型。
第Ⅱ 类:包括金皇后、巨人201、德宝、三得利、维拉和瑞西; 前期生长较快, t1出现在返青后25 d左右, 最大生长速度与旺盛生长期都处于较高的水平, 因而产量潜力最大, 可达到900~1 000 g· m-2; 属于生长快产量高的类型。
第Ⅲ 类:包括赛特一个品种, 前期生长较慢, 最大生长速度最小, 但旺盛生长期持续天数长达27.9 d, 产量潜力居中, 刈割始期在返青后55 d, 是一个生长迟缓的类型。
第Ⅳ 类:包括新疆大叶, 前期发育快, t1出现最早, 最大生长速度属中等, 但旺盛生长期最短, 产量水平最低。属于发育早而产量低的类型。
第Ⅴ 类:包括德福、甘农1号、陇东苜蓿, 前期生长发育属中等, 最大生长速度较小, 旺盛生长期属中等, 产量潜力较低。属于生长较慢产量较低的类型。
将各类的产量值进行平均, 然后用Logistic曲线进行拟合, 求得各类的平均生长曲线模型, 模型检验均达到极显著水平(图3)。
 | 图3 苜蓿不同类型的生长曲线Fig.3 Growth curve of different groups of alfalfa |
3 讨论与结论
紫花苜蓿地上生物量的动态符合Logistic生长曲线, 这点与前人的研究结果一致[9, 10, 11, 14]。但各地的回归方程差异很大。在陕西杨凌的研究结果中, 紫花苜蓿前期的缓慢生长阶段很短, t1出现在返青后2.16~9.11 d, 平均5.78 d, 远小于本项研究的26.34 d[11]; 在甘肃定西的研究表明, 缓慢生长期23~43 d, 平均33 d[9]; 在宁南山区的研究中, 缓慢生长期长达81 d, 直到6月底才进入快速生长期[10]。造成这种差异的主要原因是气候因素。在气候较温暖的地区(如陕西杨凌, 年平均气温12~14 ℃), 春季气温回升快, 植物生长迅速, 进入快速生长期早, 缓慢生长期就短。而在气候较寒冷的地区(甘肃定西, 年平均气温6.7 ℃; 宁夏西吉, 年平均气温6.0 ℃), 春季气温回升慢, 缓慢生长期就长。
不同紫花苜蓿品种生长动态的比较从总体趋势来看, 国外引进的品种生长速度快, 产草量高; 而国内品种生长速度相对较慢, 产草量也较低(北疆苜蓿例外)。这与品种的秋眠性有关, 国外品种都是半秋眠或非秋眠型, 而国内品种都是秋眠型(新疆大叶为半秋眠型)。比较生长前期缓慢生长期的变化规律, 国外品种较长, 平均26.6 d, 国内品种较短, 平均24.37 d, 这与北京地区不同秋眠苜蓿品种的生长动态相同[19], 但却与陕西杨凌不同秋眠级数苜蓿品种生物量特性[15]相反。比较各品种快速生长期的变化规律, 国内品种的较短, 平均21.18 d, 国外品种较长, 平均25.3 d, 这与上述两项研究结果都不一致。由此可见, 紫花苜蓿的生长动态是一个相当复杂的过程, 与许多因子有关, 只有把气候、土壤、施肥、灌水、品种等多种因子综合考虑, 才能得到较好的结果。这些有待于进一步深入研究。
紫花苜蓿适期刈割是栽培管理中非常重要的内容, 必须考虑其产草量、质量和持久性等因素。目前多数学者普遍认为, 苜蓿最适宜的刈割时期是早花期(即第一朵花出现至1/10花开时间), 此时刈割营养物质含量高, 再生性较好[20]。本研究提出, 根据苜蓿干物质积累的Logistic方程求出的生长速度由快速生长转变为缓慢生长的突变点(t2), 可作为刈割始期的指标。田间观察这一时期处于现蕾期, 略早于早花期。对于苜蓿刈割时期的研究报道较多。王庆锁[21]研究了苜蓿生长过程中营养成分的动态变化, 认为在现蕾末期的粗蛋白产量最高, 可以作为适宜刈割期的指标。符昕等[22]研究了苜蓿不同时期刈割对再生性的影响, 孕蕾中期刈割与初花期相比较, 株高和叶片数没有明显变化, 产草量略有下降, 但在生长后期侧枝数和节间数增加较快。说明在现蕾期刈割对于再生性没有不良影响, 而且能保证饲草质量。由此可以认为, 生长速度由快变慢的突变点(t2)作为刈割始期是合理的。这一指标在苜蓿开花很少或根本不开花的地区, 指导苜蓿生产更具有现实意义。
4 结论
不同苜蓿品种的产草量, 在第一茬其生长动态符合Logistic生长曲线。苜蓿产量潜力的高低, 不取决于最大生长速度, 也与旺盛生长期的长短无关, 而取决于它们二者的乘积。根据生长动态模型所求得的生长速度由快变慢的转折点, 可作为刈割始期, 供生产中应用。生长快而发育早的类型, 如新疆大叶, 刈割始期在5月9日; 而生长迟缓的类型, 如赛特, 刈割始期为5月22日; 其它品种居于上述品种之间。根据聚类分析结果, 第二类品种(金皇后、巨人201、德宝、三得利、维拉、瑞西)生长速度快, 产量潜力大, 从第一茬草的生长情况看, 宜作为目前生产中的推广品种。但紫花苜蓿是多年生、多次刈割的作物, 生产中还必须考虑不同年限和不同茬次的生长情况。因此, 生产中推广有待进一步验证。
参考文献
| [1] |
|
| [2] |
|
| [3] |
|
| [4] |
|
| [5] |
|
| [6] |
|
| [7] |
|
| [8] |
|
| [9] |
|
| [10] |
|
| [11] |
|
| [12] |
|
| [13] |
|
| [14] |
|
| [15] |
|
| [16] |
|
| [17] |
|
| [18] |
|
| [19] |
|
| [20] |
|
| [21] |
|
| [22] |
|