

黄河源区褐鳞苔草的固沙生物学特性
[马和, 康建军 , 赵明, 谈嫣蓉, 朱丽, 邴丹珲]
, 赵明, 谈嫣蓉, 朱丽, 邴丹珲]
, 赵明, 谈嫣蓉, 朱丽, 邴丹珲]
|
|


第一作者:马和(1962-),男,河北行唐人,副研究员,学士,研究方向为草地生态学。E-mail:[email protected]
褐鳞苔草( Carex brunnescens)是黄河源区(玛曲)沙漠化草地具有较强固沙能力的先锋植物,对维护玛曲高寒草地生态系统稳定性有重要作用。为了揭示褐鳞苔草固沙的生物学特征,本研究在野外观测了其物候学特征,地上和地下器官生长进程以及地下器官形态学特征。结果表明,褐鳞苔草返青早,生育期较长,具有二次开花和结果的生长特性。褐鳞苔草单株叶宽、单株叶长、单株叶片数量、群落密度、群落盖度和群落叶片密度从展叶期-果实成熟期-黄枯期呈显著增加而后趋于稳定;而单株叶丛高度、单株叶面积和群落叶片面积呈显著增加而后减小的趋势。褐鳞苔草地下茎(水平茎和垂直茎)和根系适宜生存的沙层深度为0-60 cm。不同沙层深度(0-20、20-40和40-60 cm)均以直径为0.2~0.5 mm的中根数量最多,在20-40 cm沙层内细根(<0.2 mm)的根系总长度均最大,0-20 cm沙层内中根根系总长度最大。在10-30 cm 沙层内,水平茎,垂直茎和主根长度、鲜重和干重均最大,40-50 cm 沙层内最小。以上结果表明,褐鳞苔草能很好地适应高寒风沙生境,可作为玛曲高寒沙漠化草地生态恢复的建群种,具有良好的推广应用前景。
Carex brunnescens is a pioneer plant that has an excellent ability to fix sand and plays an important role in maintaining the ecosystem stability of the Maqu desertified alpine grassland of the source area of the Yellow River. In this study, the biological characteristics (phenological characters, the above- and belowground growth processes, and the morphological characters of belowground organs) of C. brunnescens was studied in a field observation experiment. The results showed that C. brunnescens has the characteristics of turning green earlier than other plants, a long growth period, and two flowering and fruiting cycles. The leaf width, length, and count of individual plants, and the community plant density and leaf density showed significant increases from the leaf growth stage to the fruit maturity stage, and then tended to be stable in the withering stage, while the plant height, leaf areas of individual plants, and community leaf areas showed significant increases at first and then decreases. Underground stems (horizontal and vertical stems) and roots of C. brunnescens could survive in sand of depths 0-60 cm. There were more mid-sized roots (0.2~0.5 mm) than other root size classes in all soil layers (0-20, 20-40, and 40-60 cm). The total length of fine roots with <0.2 mm diameter was maximal in the 20-40 cm sand layer and, 0.2~0.5 mm diameter roots maximed in the 0-20 cm sand layer. The length and fresh and dry weights of the main root and horizontal and vertical stems was maximal in the 10-30 cm sand layer, and minimal in the 40-50 cm sand layer. These results showed that C. brunnescens has unique biological characteristics which could well adapt to the Maqu alpine sandy habitats, and is a species that has good potential for ecological restoration in Maqu desertified Alpine grassland.
土地荒漠化是当今人类面临的严重环境问题之一。据联合国公布的资料, 荒漠化问题已经影响到世界1/5的人口和全球2/3的国家[1, 2]。我国是世界上受荒漠化危害最为严重的国家之一, 尤其在北方草原地区, 干旱化、沙漠化日益加剧, 已经导致一系列严重的生态环境问题[3, 4]。
黄河源区(玛曲)位于青藏高原东部, 黄河自东、南、北三面环绕玛曲县, 形成“ 九曲黄河” 第一湾, 其独特的地理环境是黄河上游重要的生态屏障, 在维护黄河流域水资源和生态安全方面起着十分重要的作用[5, 6]。多年来, 在全球气候变化的大背景下, 尤其是在人为因素日益频繁的干扰和破坏下, 玛曲高寒天然草地生态环境已呈严重恶化趋势[7]。褐鳞苔草(Carex brunnescens)是莎草科苔草属多年生草本植物, 地下茎营养繁殖, 典型的无性系克隆植物, 是目前玛曲当地发现的具有强固沙能力的先锋植物, 对维护玛曲高寒草地生物多样性和生态系统稳定性有重要作用[8, 9]。褐鳞苔草可在流动沙丘上迅速繁衍, 形成固定或半固定沙地, 在一定的地形地貌条件下, 可形成少见的“ 草本植物沙堆” [10]。褐鳞苔草地下茎尤其是地下水平茎高度发达, 在生长季节, 地下水平茎不断延长, 水平茎节点上会生长垂直茎, 当地面被风沙覆盖后, 垂直茎会向上生长并迅速穿透沙面形成新的植株, 这样周而复始使地下茎形成多层网状结构起到固沙作用。当沙丘被褐鳞苔草固定以后, 其它优良牧草会迅速入侵蔓延并占据优势, 褐鳞苔草逐渐从固定沙丘退化并蔓延至其它半固定或流动沙丘上以保持种群的相对稳定[10]。到目前为止, 对褐鳞苔草的研究主要集中在种子生物学及萌发特性、种群繁殖特征以及风蚀量与种群密度、地下茎和一级根系直径的数量化关系[8, 9, 10], 而对褐鳞苔草防风固沙的生物学特性、生理生态适应性、防风固沙原理的理论研究未见报道。
本研究采用野外观测试验, 通过对玛曲高寒沙漠化草地建群植物褐鳞苔草物候观测、地上(下)器官生长进程(单株和群落)以及地下器官(地下茎和根系)形态学特征的系统研究, 探讨褐鳞苔草防风固沙的生物学特性, 进而分析其在高寒生境中所采取的适应策略, 对防治高寒草地沙漠化以及促进退化草地生态恢复具有重要的实践意义。
试验地点位于甘肃省甘南州玛曲县欧拉乡黄河沿岸(100° 45'45″-102° 29'00″ E, 33° 06'30″-34° 30'15″ N), 具有明显的高原大陆性气候特点, 海拔为3 300~4 806 m, 立地区划亦属“ 黄河一级阶地区” [11]。一年中无明显的四季之分, 年平均气温 1.1 ℃, 年平均降水量615.5 mm, 年均蒸发量1 353.4 mm。全县地貌由高山、山地、丘陵和河岸阶地构成, 土壤类型以暗棕壤、高山草甸土、亚高山草甸土为主[12]。试验区位于玛曲县欧拉乡欧强村黄河沿岸的活动沙丘, 距县城5 km, 面积为9.8 hm2, 地形为河岸阶地, 土壤为沙质土, 主要植物为褐鳞苔草、粗壮嵩草(Kobresia robust)、赖草(Leymus secalinus)及垂穗披碱草(Elymus nutans)等。
1.2.1 物候观测 观测内容及方法参照中国科学院地理科学与资源研究所制定的草本植物物候观察法[13]。
1.2.2 地上器官生长进程观测(单株和群落)
观测内容:褐鳞苔草地上器官在不同生育期(展叶期、开花盛期、果实成熟期及黄枯期)的生长速度。单株生长进程主要测定叶片数量(枚)、叶宽度(最宽处)(mm)、叶长度(mm)、叶丛盖度(自然高度mm)和叶面积(mm2)等。群落生长进程主要观测群落密度(株· m-2)、群落盖度(投影盖度, %)、群落叶片密度(枚· m-2)及群落叶片面积(cm2· m-2)。
观测方法:在固定沙地、半固定沙丘及流动沙丘的风蚀区选择代表性样地, 在褐鳞苔草生育期观测单株和群落生长进程。单株(地上可分单株)生长进程实行定株观测(采用样方内植株), 观测株数20株以上(大样本)。群落生长进程实行固定样方观测, 样方大小1 m× 1 m, 样方数量20以上(小样本), 样边界做固定标志。叶面积(mm2)和群落盖度计算采用Webcam物品颜色分析系统。
1.2.3 地下器官形态学特征观测 地下茎观测:分别在半固定沙丘和流动沙丘的风蚀区选取代表性样地, 样地重复3次, 从上到下挖出整个褐鳞苔草剖面结构, 深度2.0 m, 宽度25 cm, 横切面厚度5 cm。依次记录每20 cm厚度内横切面水平茎、垂直茎个数、成活数、茎节发芽数。
根系观测方法与地下茎相同。在半固定沙丘的风蚀区和流动沙丘的风蚀区分别设立固定样地(1 m× 1 m), 样地重复3次。采用分层分段挖掘法对褐鳞苔草根系分布状况进行调查。按土壤层次(0-20、20-40及40-60 cm)挖出全部地下部分后, 将地下茎和根系分离, 除去土壤后, 仔细挑拣出全部活根, 分别按细根(< 0.2 mm)、中根(0.2~0.5 mm)和粗根(> 0.5 mm)分层计数根数; 并清洗根表面, 用游标卡尺逐一测量根系粗度; 用卷尺测量各级的根长度, 计算一级根系总长度。
1.2.4 地下器官生长进程(地下茎和一级根系)
观测内容:褐鳞苔草地下器官在不同生育期(展叶期、开花盛期、果实成熟期及黄枯期)的生长速度和生长期结束后不同沙层深度生物量的变化。
生长速度观测:主根长度, 主根直径, 地下茎(水平茎和垂直茎)长度、分枝数和粗度。
观测方法:在半固定沙丘和流动沙丘的风蚀区选取代表性样地, 样地重复3次, 每个样地固定3~5株单株或地下主茎在生育期内定期观测地下器官的生长速度。在地下器官分布层设垂直剖面或反倾斜剖面, 使根系和地下茎沿剖面生长便于观测, 剖面用大孔径尼龙网加透明板材固定保护。第1次观测在根系或地下茎固定位置拴标记绳或在透明板材上做标记, 以后测量标记部位到先端部位的长度。
生物量观测内容:地下茎和根系的数量指标(长度、鲜重及干重等)。
观测方法:在地形地貌和周边环境相同(主要是沙丘形状、大小和部位)的情况下, 分别在半固定沙丘和流动沙丘的风蚀区选取代表性群落样方(面积为1 m× 1 m), 样地重复3次, 按土壤层次(0-10、10-20、20-30、30-40及40-50 cm)挖出全部地下器官后, 将地下茎和根系(一级)分离, 挑拣出全部活茎和根, 用钢卷尺测量地下茎和根系长度, 计算各沙层地下茎和根系总长度。然后用蒸馏水快速冲洗地下茎和根系表面, 用吸水纸吸干表面水分, 称鲜重; 然后将鲜材料80 ℃烘干至恒重后, 称干重。
褐鳞苔草单株叶片数量、叶宽、叶长、叶丛盖度、叶面积和群落密度、盖度及叶片密度、叶片面积, 褐鳞苔草剖面形态学特征(横切面水平茎、垂直茎个数、成活数、茎节发芽数, 根系粗度和长度), 地下器官生长特征(主根长度、直径及生物量, 地下茎直径、长度、分枝数及生物量等)等观测数据用SPSS 15.0 (SPSS Inc., USA)软件进行统计分析, 用Excel作图。
褐鳞苔草萌动期较早(4月中旬), 黄枯期较迟(9月上旬), 生育期较长(5个月)(表1); 褐鳞苔草进入萌动期后, 首先水平茎和垂直茎优先生长, 同时伴随着地下芽和垂直茎的形成, 随后植株开始出土, 展叶, 迅速进入开花期(持续45 d); 具有二次开花和结果, 且果实成熟期较早的生物学特性, 果期持续时间长(3个月), 种子成熟后开始脱落, 散布, 随后植株进入黄枯期。
| 表1 褐鳞苔草物候观察记录表 Table 1 Phenological observations of Carex brunnescens |
在固定沙地、半固定及流动沙丘的风蚀区, 褐鳞苔草单株叶宽度、叶长度和叶片数量及群落密度、群落盖度和群落叶片密度从展叶期开始到开花盛期、果实成熟期呈现显著增加(P< 0.05)(表2、表3), 到黄枯期后呈趋于稳定的趋势, 而单株叶丛高度、叶面积和群落叶片面积从展叶期开始到开花盛期、果实成熟期显著增加然后到黄枯期显著减小(表2、表3)。
| 表2 褐鳞苔草单株生长进程记录表 Table 2 Single plant growth process of Carex brunnescens |
| 表3 褐鳞苔草群落生长进程记录表 Table 3 Community growth process of Carex brunnescens |
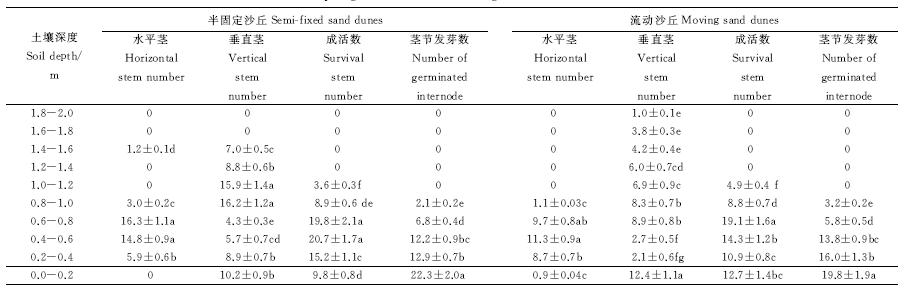
2.3.1 褐鳞苔草地下茎形态学特征观测 在半固定沙丘的风蚀区和流动沙丘的风蚀区, 从上往下挖至2.0 m深将褐鳞苔草剖面完整挖出以后, 发现深度在1.0 m以下时褐鳞苔草水平茎很少; 深度在1.6-2.0 m时, 垂直茎很少; 深度0-1.0 m时, 水平茎和垂直茎均能够成活, 并且深度在0-0.6 m时, 水平茎和垂直茎均能够发芽长出地下茎; 深度在0-0.2 m时, 水平茎和垂直茎上茎节发芽数量均最多, 0.2-0.4 m时次之, 1.0-2.0 m时, 茎节发芽数很少(表4)。
| 表4 褐鳞台草地下茎剖面形态学特征记录表 Table 4 Profile morphological characteristics of belowground rhizomes of Carex brunnescens |
2.3.2 褐鳞苔草根系形态学特征观测 在半固定沙丘和流动沙丘的风蚀区, 不同深度沙层(0-20、20-40及40-60 cm)内, 褐鳞苔草根系总数、一级根系总长度和粗度呈显著变化的趋势(表5)。根系数量方面, 不同沙层深度均以直径为0.2~0.5 mm的中根数量最多。直径> 0.5 mm的粗根数量在0-20和40-60 cm土层内较多, 且差异不显著(P> 0.05); 在20-40 cm土层内最少。直径< 0.2 mm的细根数量在20-40和40-60 cm土层内较多, 0-20 cm土层内最少。根系长度方面, 20-40 cm土层内细根根系总长度最大, 40-60 cm土层内粗根总长度最大, 0-20 cm土层内中根系总长度最大。
| 表5 褐鳞台草根系形态学特征性记录表 Table 5 Root morphological characteristics of Carex brunnescens |
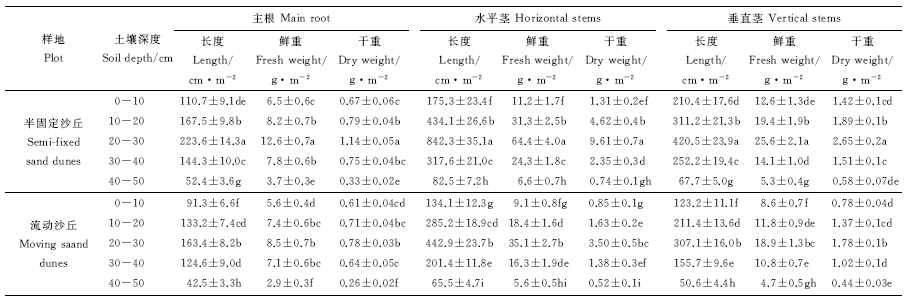
在半固定和流动沙丘的风蚀区, 褐鳞苔草地下器官在不同生长时期的生长速率和生长期结束后不同沙层深度(0-10、10-20、20-30、30-40及40-50 cm)的生物量呈显著变化的趋势(P< 0.05)。主根、水平茎和垂直茎长度、水平茎分枝数从展叶期开始到开花盛期、果实成熟期生长不断加快, 而到黄枯期后呈减小的趋势, 而主根直径、水平茎和垂直茎直径的变化差异不显著(P> 0.05); 主根长度、水平茎和垂直茎长度、分枝数在开花盛期和果实成熟期生长速率最快(表6)。水平茎、垂直茎和主根主要分布在10-30 cm沙层内, 并且10-30 cm沙层内水平茎、垂直茎和主根长度、鲜重和干重均最大; 30-40 cm沙层内次之, 40-50 cm沙层内最小(表7)。
| 表6 地下器官生长进程记录表 Table 6 Growth process of underground organs of Carex brunnescens |
| 表7 褐鳞台草地下器官生物量变化记录表 Table 7 Biomass of underground organs of Carex brunnescens |
荒(沙)漠化是植被覆盖退化、风蚀加剧的过程, 适应流沙环境植物的快速繁衍能够逆转荒漠化(沙漠化)过程, 并能够固定流沙。水分亏缺和养分贫瘠是沙漠化生境的基本特征, 植物必须经受养分匮乏和干旱胁迫的双重考验, 才能维持其存活与繁衍[14]。苔草属(Carex)植物是莎草科(Cyperaceae)多年生草本, 地下茎营养繁殖, 属于典型的根茎型克隆植物, 苔草属植物生态幅很宽, 数量较多, 具有很强的抗逆性特征(抗寒、抗旱、耐践踏及抗风蚀沙埋等)[10, 15, 16]。褐鳞苔草属于典型的根茎型克隆植物, 是目前玛曲当地沙漠化草地生态系统发现的重要建群种和优势种。克隆植物在植物界广泛存在, 生理整合或克隆整合是克隆植物所共同具有的重要适应性特征之一, 同时也是其适应异质性生态环境的重要策略之一[17]。然而, 克隆植物的生理整合也并非是普遍存在的, 对于某些植物种类来说, 相连克隆分株之间只往往存在着程度很低的生理整合, 甚至不存在生理整合[18]。观测发现, 褐鳞苔草具有发达的地下结构, 埋藏于流沙下50 cm深度时其地下茎仍具有萌发和生长到地面的能力, 并且褐鳞苔草相连克隆分株之间存在着程度很高的生理整合作用。植株产生细长的下伸和横走的根茎, 其地下根茎分枝类型为单轴型, 可以产生很多的根茎顶端芽和根茎节芽向上发育形成新的分株, 从而实现基株的空间扩展和分株种群的持续更新, 使得褐鳞苔草能够通过克隆生长, 产生与母株相连的新的子代分株, 形成克隆[10]。相对于母株来说, 新的分株通过空间上的移动, 实现褐鳞苔草在水平空间的拓展, 进入到不同的沙漠生境斑块。
植物的生长规律受自身生物学特性和生态环境条件的双重制约[19]。植物的生长主要表现在地上和地下部分生物量的增加, 还表现为地上部分高度的增长和地下部分深度的增加[19, 20]。受外界逆境环境因子的影响, 植物地上和地下部分生长动态过程往往具有明显的季节性变化[21, 22, 23, 24]。植物地下部分在适宜的温度、水分和养分供给条件下生长较快, 增加垂直根系的生长可以显著增加植物对水分的吸收和利用[24, 25], 而且细根可承担植株的吸收功能, 反映根系的吸收能力[26, 27]; 同时可以有效利用深层土壤水, 从而保证植物在逆境环境下维持其正常的生理活动[28]。从植物的生长动态过程来看, 地下部分的生长优势就是植物适应逆境环境的生长策略和重要特征[19]。在长期适应各种逆境环境的过程中, 植物通过平衡各种性状、优化资源配置, 在不同的环境中形成不同的适应策略, 最终适应复杂不变的环境。植物资源分配格局反映了植物发育对逆境的响应规律和适应对策, 将更多资源投资于地下部分是对逆境环境的一种适应性策略[14, 29]。对沙生植物而言, 为了更有效地利用环境与资源, 沙生植物在进入生长期后营养生长速度很快, 往往在较短的时间内即可完成高生长和叶片的生长, 很快进入花期、结果的繁殖期。沙生植物以较短的营养生长期换取较长的繁殖生长期, 是植物对沙漠环境不可预测性的一种适应, 这种对策可以使植物在生长后期遭遇不良环境时迅速完成繁育任务, 避免恶劣环境下种群遗传基因的丢失。本研究表明, 为了使遗传基因完成每年的传递任务, 褐鳞苔草在返青后采取了花叶同时生长的适应性策略, 并且具有二次开花和结果的适应性特征, 这种一生多次繁殖的策略, 使得褐鳞苔草每年都有繁殖的机会, 可以产生繁殖体, 即使在不可预知的风沙环境中也可以更多地保留遗传基因, 使得其在风蚀和沙埋环境中具有了很强的适应能力。在沙漠生境中, 不同植物种类地上不同器官的生长发育速率存在较大差别, 对于褐鳞苔草而言, 茎干的发育往往较快, 在较短时间内达到正常高度, 这样不仅能够较快地占据生长空间, 增加竞争能力, 而且为叶片的扩展提供有利条件。除此之外, 褐鳞苔草地下器官的生长速度都较地上部分快得多。当其地上部分刚刚萌动时, 其地下部分常常深入地下数厘米至数十厘米。而且, 在年度生长过程中, 褐鳞苔草也总是将相对更多的能量、水分和养分储存于地下部分供其生长发育, 从而具有了庞大的地下储存器官和较高较强的适应能力。总之, 植物的环境适应性是植物对环境长期适应的综合遗传特性, 不仅与植物的解剖结构有关, 还取决于植物的外部形态特征及内部的生理生化特性等方面[30, 31]。Bradshaw和Hardwick[32]认为, “ 进化是植物对外界环境压力几乎不可避免的结果” 。每一种植物都形成了其适应逆境环境时的复杂生存机制, 以确保其能够在特定的环境中生存、繁衍和发展[33]。然而, 尽管褐鳞苔草繁殖和适应能力极强, 但其种子的萌发受到系统发育约束力的严重控制, 致使种子萌发率很低[8, 9], 很难应用于固沙实践。在高寒生境下, 或许通过探讨苔草属植物种子繁殖特性与系统发育的相互关系将帮助揭示其种子休眠率高的原因。目前, 高寒沙漠化草地通过特定草本植物固沙能力的研究很少, 特别是对褐鳞苔草生态生理适应性、防风固沙原理以及沙包形成过程及必备条件的理论研究还未见报道, 而且受到研究方法和技术手段的限制, 褐鳞苔草对高寒环境适应特征的系统研究始终存在着局限和不足, 进而也影响到通过地上/地下部分的生长关系来分析认识其对环境的适应性规律。到目前为止, 陆地生态系统的大量研究主要涉及植物的地上部分, 地下部分的研究还相对甚少, 可以通过对玛曲高寒沙漠化草地褐鳞苔草地上/地下部分生长动态的观测研究, 分析其生长的动态变化规律, 进而探讨其在高寒风沙生境中所采取的适应性策略。
The authors have declared that no competing interests exist.
| [1] |
|
| [2] |
|
| [3] |
|
| [4] |
|
| [5] |
|
| [6] |
|
| [7] |
|
| [8] |
|
| [9] |
|
| [10] |
|
| [11] |
|
| [12] |
|
| [13] |
|
| [14] |
|
| [15] |
|
| [16] |
|
| [17] |
|
| [18] |
|
| [19] |
|
| [20] |
|
| [21] |
|
| [22] |
|
| [23] |
|
| [24] |
|
| [25] |
|
| [26] |
|
| [27] |
|
| [28] |
|
| [29] |
|
| [30] |
|
| [31] |
|
| [32] |
|
| [33] |
|
黄河源区褐鳞苔草的固沙生物学特性
马和, 康建军, 赵明, 谈嫣蓉, 朱丽, 邴丹珲
Biological sand-fixation characteristics of Carex brunnescens in Yellow River source region
Ma He, Kang Jian-jun, Zhao Ming, Tan Yan-rong, Zhu Li, Bing Dan-hui