


{kind=link}
{kind=link}
{kind=link}
{kind=link}
青海湖东沙化治理过程中土壤酶活性及养分含量特征
[魏小星 ]
]
]
|
|


本研究以青海湖东沙化治理区建植5年所形成的披碱草( Elymus dahuricus)、碱茅( Puccinellia distans)和早熟禾( Poa annua)栽培草地为研究对象,通过测定土壤有机质、氮、磷、钾养分含量以及脲酶、过氧化氢酶和转化酶活性,探讨了在高寒干旱沙化生态系统中不同草本植物作用下土壤养分和土壤酶的变化特征及其相互关系。结果表明,1)人工治沙植被作用下,土壤养分和土壤酶活性均显著提高( P<0.05);2)早熟禾在青海湖区域治沙过程中对土壤有机质、碱解氮、速效磷、全钾和速效钾的改良效果显著( P<0.05);3)在3种栽培草地处理下土壤脲酶活性在表层土壤中普遍显著高于流动沙丘对照( P<0.05),披碱草和早熟禾栽培草地的脲酶活性较高,0-20 cm土层中土壤过氧化氢酶活性在3个栽培草地中均显著高于对照( P<0.05),且早熟禾栽培草地中土壤过氧化氢酶活性最高,土壤转化酶在各栽培草地间差异不显著( P>0.05);4)各栽培草地中土壤脲酶和过氧化氢酶普遍与土壤中的碳氮磷养分存在显著相关性( P<0.05),可以作为沙化治理过程中评价土壤恢复的有效指标。
This study focused on the artificial grasslands established with three different plant species Elymus dahuricus (Ely), Puccinellia distans (Puc) and Poa annua (Poa) for five years to control desertification to the east of Qinghai Lake. To assess the effects of these three artificial grasslands on sand soil nutrient content, soil enzyme activities and their relationships, we analysed soil nutrient (nitrogen, phosphorous, and potassium) and organic matter contents, as well as the activities of three soil enzymes, namely, urease, catalase, and invertase. The results showed that: 1) soil nutrient and soil enzyme activity were significantly improved after the grasses were planted. 2) Compared with the other two treatments, Poa had significant effects on improving soil organic matter, available nitrogen, available phosphorus, total potassium, and available potassium. The soil nutrient content had a significant positive relationship with soil urease activity. 3) Soil urease activity was much greater in the Ely and Poa treatments, and the activities in all the three treatments were higher than the control in the surface soil layer. In the 0-20 cm soil layer, the activities of soil catalase in the three treatments were higher than that in the control, with Poa showing the highest activity. All three treatments showed no significant effect on the activity of soil invertase. 4) The activities of soil urease and catalase both had a general significant relationship with soil nutrient content, which could be taken as the effective index for the measurement of desertification control process.
气候变化和人类活动等诸多因素引起的干旱地区土地沙化, 已经成为全球关注的生态环境热点问题[1, 2]。青海省地处我国沙漠化面积最大、海拔最高的沙漠地带, 沙化面积约占全省国土面积的16.6%, 且在不断扩大[3, 4]。1956-2000年间青海湖流域沙漠化面积以每年18.1 km2的速度增加, 2005-2006年移动沙丘以每年23.9 m的速度移动, 青海湖流域已成为青海省沙化治理的重点地区[5]。
人工固沙草地在建植3~5年后, 植物-土壤的补偿性生长与反馈形成固沙草地初级生态系统, 其防风固沙的核心是植物-土壤界面的反馈与补偿作用驱动地面植物群落的进化以及沙质土壤的改良[6]。土壤酶作为土壤的组成部分, 在有机质转化和养分循环中起重要作用, 与土壤理化性质息息相关, 不仅能反映土壤中生物代谢和物质转化情况, 还能较好地反映土壤的表观肥力[7, 8]。土壤养分是沙化治理过程中植物生长的基础, 植物的生存以及生长发育所需的养分很大程度上依赖于酶促反应分解的产物。土壤酶在干旱贫瘠土壤中具有很强的抗逆性, 可在不利于作物生长的逆境条件下调节土壤养分转化[9, 10, 11, 12, 13, 14]。近几年, 对不同生境及不同干扰下土壤酶的活性及影响因素研究很多, 其中包括沙地、荒漠、草原、山地、丘陵、滨海湿地和水陆交错带等不同生境[15, 16, 17, 18, 19, 20, 21], 以及施肥、轮作、免耕、退耕、秸秆还田、补播、微灌和刈割等农艺措施[22, 23, 24, 25, 26, 27, 28, 29], 还有诸如演替、氮沉降和增温等生态过程[30]。总体来说, 在沙化治理过程中土壤酶的研究较少, 在沙化恢复过程中土壤酶和养分的特征变化及土壤养分的形成与土壤酶之间的关系仍然不是很明确。
本研究以青海湖东沙化治理区建植5年所形成的披碱草(Elymus dahuricus)、碱茅(Puccinellia distans)和早熟禾(Poa annua)栽培草地为研究对象, 测定人工治沙植被下土壤酶活性以及土壤理化性质的变化, 以期揭示沙化地区栽培草地生态系统中植物-土壤界面中土壤酶、土壤理化性质受植物的响应及反馈作用, 为进一步研究综合、科学的固沙技术提供理论依据。
试验地位于青海省海南州共和县青海湖湖东, 地理坐标为30° 01'46″ N、100° 19'46″ E, 海拔3 010 m, 风向以北风和西北风为主, 风速多在2~3 m· s-1, 年降水量为300 mm 左右, 年平均蒸发量1 100 mm左右, 原生植被平均盖度为5%~6%。
2008年在青海湖东流动沙丘试验地上设置六角形空心砖(直径25 cm)沙障条带, 沙障条带高度30 cm、间距100 cm、东西走向(与试验区风向垂直); 在沙障条带内种植披碱草(Ely处理)、碱茅(Puc处理)、早熟禾(Poa处理), 种植时间为当年6月。每个处理面积1.2 hm2, 播量(Ely:45 kg· hm-2, Puc:15 kg· hm-2, Poa:20 kg· hm-2), 条播, 施有机肥400 kg· hm-2、磷酸二铵100 kg· hm-2作为基肥, 播种深度2.5 cm。试验区采取了严格的禁牧和管理措施, 2013年将试验区内每个处理设置6个重复小区, 每个小区面积0.2 hm2。对照区(CK)是自2008 年以来就禁牧的流动沙丘。
2013-2015年连续3年对试验区沙障内Ely、Puc、Poa处理及CK分层采集0-10、10-20和20-30 cm的土壤样品, 采集时间为每年9月份, 采用多点混合采样, 每个处理6次重复。土样用自封袋带回实验室风干, 过0.2 mm后用于土壤养分和酶活性测定。
采用半微量凯氏定氮法测定全氮, 钼锑抗比色法测定全磷, 酸溶法(HF-HClO4)测定全钾, 碱解扩散法测定碱解氮, 碳酸氢钠浸提-钼锑抗比色法测定速效磷, 乙酸铵浸提法测定速效钾, 重铬酸钾-外加热法测定土壤有机质[31]。
| 表1 2013-2015年试验地植被情况 Table 1 Plant characteristics in experimental field from 2013 to 2015 |
过氧化氢酶采用滴定法测定, 其活性以单位土重的30 min内消耗的高锰酸钾升数表示。土壤脲酶采用扩散法测定, 其活性以1 g土壤在37 ℃培养15 h释放的NH3-N的毫克数表示。转化酶采用滴定法测定, 其活性以1 g土壤的硫代硫酸钠毫升数表示。
使用SPSS 17.0方差分析, 用Tukey法进行多重比较(P< 0.05)。使用SigmaPlot 12.0进行作图 。
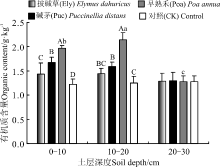
2.1.1 土壤有机质含量对栽培草地处理的响应 在0-10 cm土层, 3个栽培草地处理有机质含量表现为Poa> Puc> Ely> CK, 且差异达到显著水平(P< 0.05), Poa处理有机质含量最高, 达到1.97 g· kg-1; 10-20 cm土层, 3个处理的土壤有机质含量高于对照, 其中Poa处理的土壤有机质含量最高, 达到了2.02 g· kg-1, 显著高于Puc和Ely处理(P< 0.05); 20-30 cm土层, 各处理有机质含量与对照差异不显著(P> 0.05)(图1)。
Ely和Puc处理有机质含量随着土层深度增加逐渐降低, 但差异不显著(P> 0.05); Poa处理下有机质含量随着土层深度的增加呈先升高再降低的趋势, 差异显著(P< 0.05)(图1)。
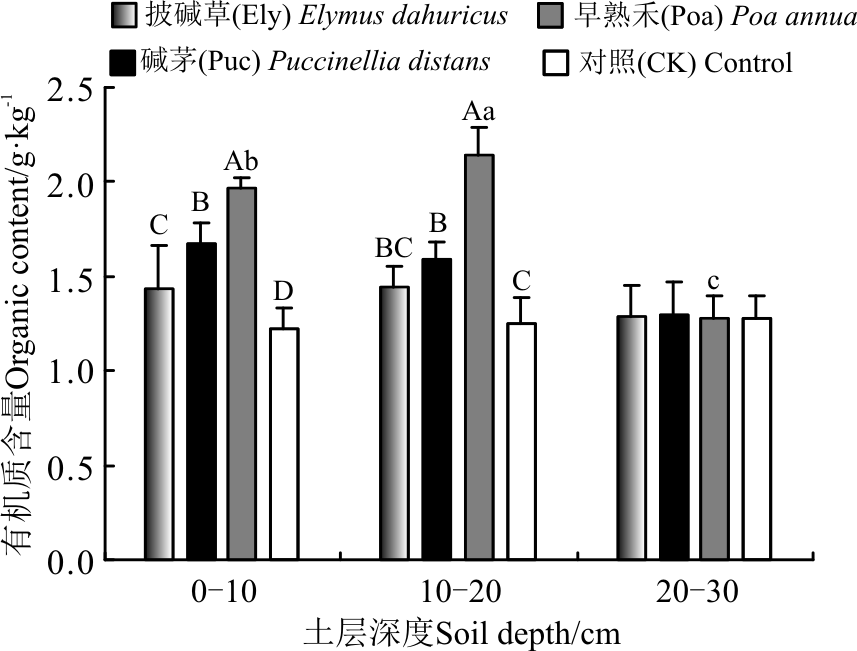
2.1.2 土壤氮素含量对栽培草地处理的响应 在0-10 cm土层, 3个栽培草地处理土壤中全氮含量均显著高于对照(P< 0.05), 3个处理全氮含量表现为Ely> Poa> Puc, 3个处理碱解氮含量表现为Poa> Puc> Ely, 其中Poa处理碱解氮含量显著高于Ely、Puc处理及对照(P< 0.05), 达到62.54 mg· kg-1; 在10-20 cm土层, 3个栽培草地处理土壤全氮含量表现为Puc和Poa显著高于Ely和对照(P< 0.05), 碱解氮含量各处理间均差异不显著(P> 0.05); 20-30 cm土层, 3个处理全氮和碱解氮含量均与对照差异不显著(P> 0.05)(图2)。
 | 图1 植被类型对土壤有机质含量的影响注:图中不同大写字母表示同一土层不同处理间差异显著(P< 0.05), 不同小写字母表示同一处理不同土层间差异显著(P< 0.05)。图2、图3和图4同。Fig.1 Effect of vegetation type on soil organic matter contentNote: Different capital letters in the same soil layer indicate significant difference in different treatments at the 0.05 level, whereas different lowercase letters indicate in the same treatment indicate significant difference amon different soil layers at the 0.05 level; similarly for Table 2, Table 3, and Table 4. |
3个栽培草地处理间全氮含量在各土层变化不显著(P> 0.05); Ely和Puc处理碱解氮含量随着土层深度的增加呈先升高再降低的趋势(P< 0.05), Poa处理土壤碱解氮含量随土壤深度的增加逐渐降低的趋势(P< 0.05)(图2)。
 | 图2 植被类型对土壤全氮和碱解氮含量的影响Fig.2 Effect of vegetation type on the amounts of total nitrogen and available nitrogen in soil |
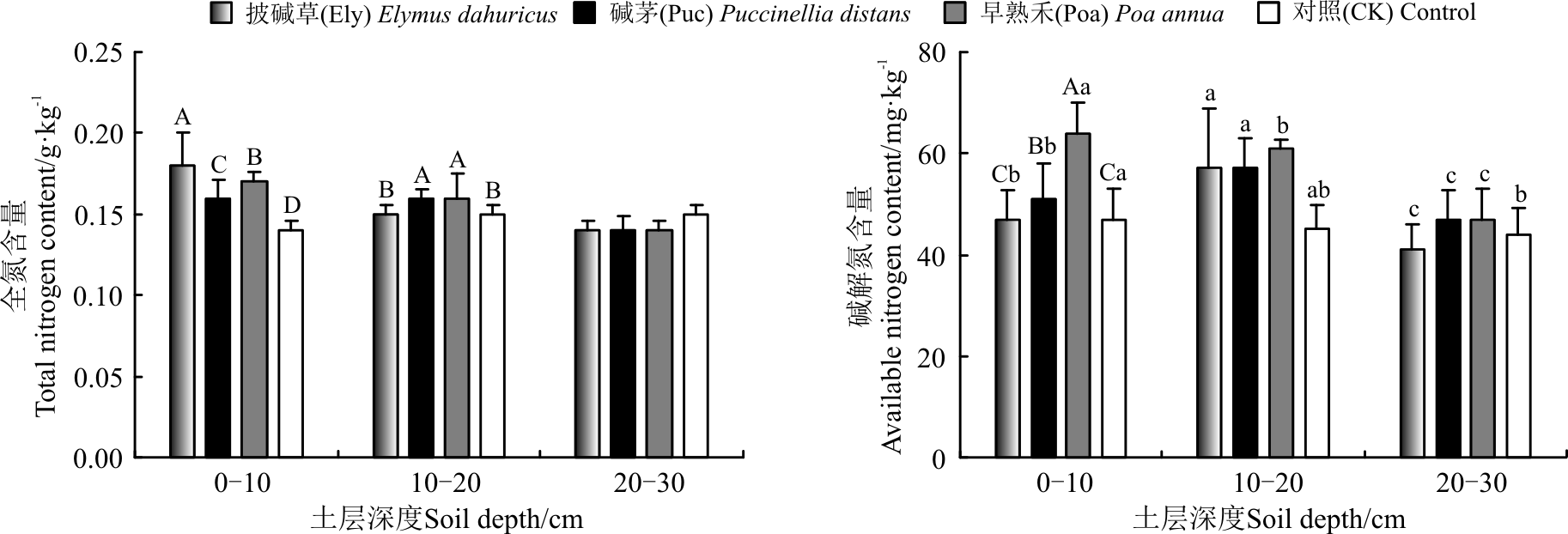
2.1.3 土壤磷素含量对栽培草地处理的响应 不同植被类型下, 土壤全磷含量差异不显著(P> 0.05); 在0-10、10-20 cm土层, 3个栽培草地处理土壤速效P含量均显著高于对照(P< 0.05), 3个处理间速效磷含量Poa> Puc> Ely, 其中Poa处理在0-10 cm土层速效磷含量达到2.95 mg· kg-1, 在10-20 cm土层达到4.89 mg· kg-1, 显著高于对照(P< 0.05)(图3)。
 | 图3 植被类型对土壤全磷和速效磷含量的影响Fig.3 Effect of vegetation type on total and available phosphorus amounts in soil |
3个栽培草地处理土壤全磷含量在各土层差异不显著(P> 0.05); 3个处理速效磷含量随着土层深度的增加呈先升高再降低趋势, 在10-20 cm土层中达到最大(P< 0.05)(图3)。
2.1.4 土壤钾元素含量对栽培草地处理的响应 3个栽培草地处理下土壤全钾含量差异不显著(P> 0.05); 3个处理速效钾含量在0-10、10-20 cm土层均显著高于对照(P< 0.05), 3个处理间速效钾含量表现为Poa> Puc> Ely; 在20-30 cm土层Poa处理速效钾含量显著高于其它处理(P< 0.05), 达到51.54 mg· kg-1(图4)。
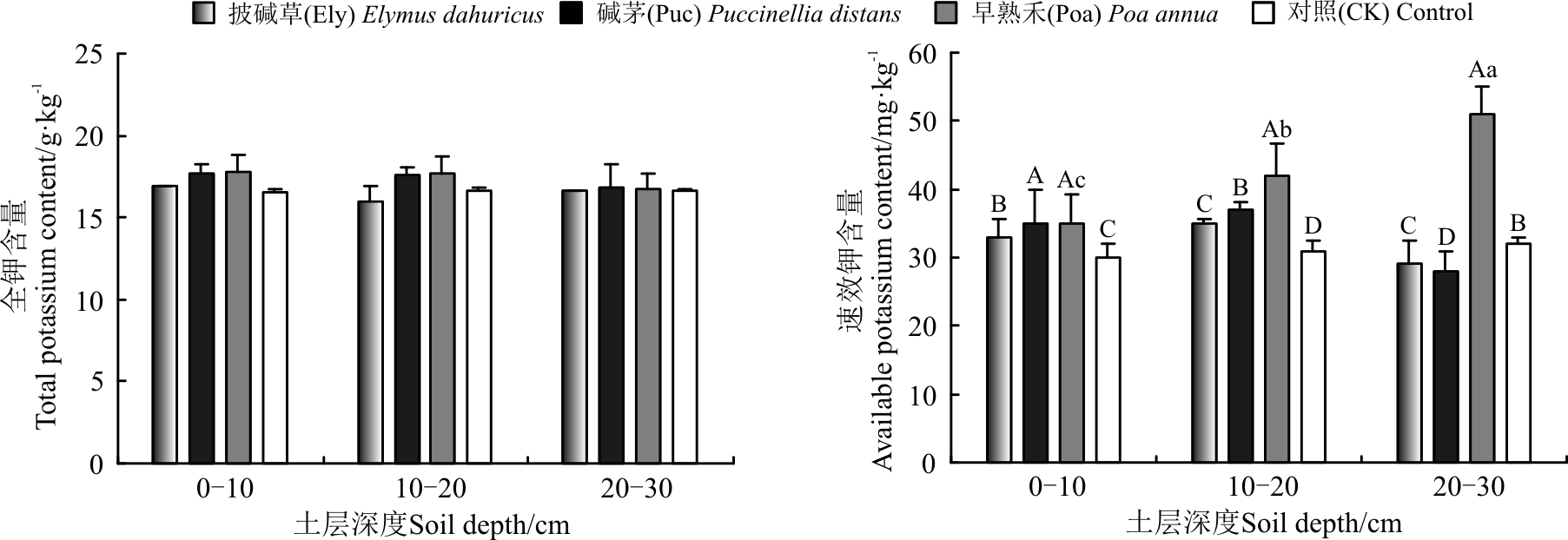 | 图4 植被类型对土壤全钾和速效钾含量的影响Fig.4 Effect of vegetation type on amounts of total and available potassium in soil |
3个栽培草地处理间全钾含量在各土层差异不显著(P> 0.05); 速效钾含量在Ely、Puc处理3个土层深度差异不显著(P> 0.05), Poa处理随土层深度的增加呈升高趋势(P< 0.05)(图4)。
2.2.1 栽培草地处理对土壤脲酶活性的影响 不同土层土壤脲酶活性对处理均具有明显的响应。具体表现为, 2013年, 0-10 cm土层土壤脲酶活性在Ely和Poa处理下均显著高于CK(P< 0.05)(表2), 10-20 cm土层土壤脲酶活性在Ely、Puc和Poa处理下均显著高于CK(P< 0.05), 20-30 cm土层土壤脲酶活性只有在Poa处理下显著高于CK(P< 0.05)。2014年, 0-10和10-20 cm土层土壤脲酶活性Poa处理显著高于CK(P< 0.05)。2015年, 0-30 cm各土层均表现为3个处理显著高于CK(P< 0.05)。
2.2.2 不同栽培草地处理下土壤过氧化氢酶活性变化特征 2013年, 0-10和10-20 cm土层土壤过氧化氢酶活性在3个处理下均显著高于CK(P< 0.05)(表2), 且3个处理间Poa 处理下土壤过氧化氢酶活性最高; 20-30 cm土层, 土壤过氧化氢酶活性在不同处理之间差异均不显著(P> 0.05)。2014年, 0-10 cm土层土壤过氧化氢酶活性3个处理均显著高于CK(P< 0.05); 10-20 cm土层, 土壤过氧化氢酶活性显著高于CK(P< 0.05)。2015年, 0-20 cm土层均为3个处理显著高于CK(P< 0.05)。
| 表2 2013-2015年土壤脲酶活性、过氧化氢酶活性和转化酶活性比较 Table 2 Different activities of urease, catalase and invertase in soil from 2013 to 2015 |
2.2.3 不同栽培草地处理下土壤转化酶活性变化特征 3年的结果均显示, 0-30 cm土层中, Poa、Ely和Puc这3个处理对土壤转化酶没有明显影响(P> 0.05)(表2); 3个处理中Puc和Ely对土壤转化酶影响最大, 但差异不显著(P> 0.05)。
为了探讨不同栽培草地处理土壤酶活性与土壤养分各因子之间的关系, 本研究对二者进行了相关性分析(表3-表5)。Ely处理下, 在0-10 cm土层, 土壤脲酶与土壤各养分间相关性不显著(P> 0.05); 10-20 cm土层, 土壤脲酶与土壤全磷和全钾含量显著正相关(P< 0.05); 20-30 cm土层, 土壤脲酶与土壤有机质极显著正相关(P< 0.01), 与土壤碱解氮显著正相关(P< 0.05), 与全磷显著负相关。在0-10 cm土层过氧化氢酶与土壤速效磷极显著正相关(P< 0.01), 与土壤速效钾显著正相关(P< 0.05)。转化酶在0-10 cm土层与碱解氮, 在20-30 cm土层与土壤有机质呈显著正相关(P< 0.05)(表3)。
| 表3 披碱草处理土壤酶活性与养分之间的相关系数 Table 3 Correlation coefficients for enzyme activity in soil with Elymus dahuricus experiment and soil nutrient |
Puc处理下(表4), 0-20 cm土层, 土壤脲酶与土壤各养分之间无显著相关性(P> 0.05), 在20-30 cm土层, 土壤脲酶与土壤有机质和全钾极显著正相关(P< 0.01), 与土壤碱解氮显著正相关(P< 0.05), 与速效钾显著负相关(P< 0.05)。0-10 cm土层, 土壤过氧化氢酶与速效磷和速效钾极显著正相关(P< 0.01); 10-20 cm土层, 土壤过氧化氢酶与土壤全钾极显著负相关(P< 0.01); 20-30 cm土层, 其与土壤速效磷显著负相关(P< 0.05)。土壤转化酶除在10-20 cm土层与土壤碱解氮显著正相关(P< 0.05), 在20-30 cm土层除与土壤有机质显著正相关外, 其它均无显著性(P> 0.05)。
| 表4 碱茅处理土壤酶活性与养分之间的相关系数 Table 4 Correlation coefficients for enzyme activity in soil with Puccinellia distance experiment and soil nutrients |
Poa处理下(表5), 土壤脲酶与土壤碱解氮在0-10 cm土层显著正相关(P< 0.05), 与速效钾显著负相关(P< 0.05); 在10-20 cm土层, 脲酶与碱解氮和全磷极显著正相关(P< 0.01), 与全氮显著正相关(P< 0.05), 与速效磷极显著负相关(P< 0.01); 在20-30 cm土层, 脲酶与土壤有机质、碱解氮、全钾和速效钾极显著正相关(P< 0.01)。过氧化氢酶在0-10 cm土层与土壤速效磷极显著正相关(P< 0.01), 与土壤全钾显著负相关(P< 0.05); 10-20 cm土层除与土壤全磷显著正相关(P< 0.05), 与土壤速效磷极显著负相关(P< 0.01)。土壤转化酶除在0-10和20-30 cm土层与碱解氮分别极显著(P< 0.01)和显著正相关(P< 0.05), 在10-20 cm土层与土壤有机质和全钾显著正相关(P< 0.05)外, 与其它均无显著相关性(P> 0.05)。
| 表5 早熟禾处理土壤酶活性与养分之间的相关系数 Table 5 Correlation coefficients for enzyme activity in soil with Poa annua experiment and soil nutrients |
设置沙障能够在流动沙丘中建植披碱草、碱茅、早熟禾栽培草地并形成稳定的生态系统, 草地固沙生态系统稳定持续的核心是植物界面-土壤界面的反馈与补偿作用驱动地面植物的进化以及沙质土壤的改良[32, 33, 34, 35, 36, 37]。不同草本植物的种植影响着土壤有机质的生产, 土壤有机质与全氮、速效磷、速效钾含量等密切相关, 有机质含量及其存在状况深刻地影响着土壤理化性质及生物学性质[38, 39]。本研究与以往固沙植被对沙化土壤理化性质的改善或类似生境的研究结果是一致的, 在利用红柳(Tamarix ramosissima)对川西北沙化草地进行生态修复过程中, 随着修复年限的增加, 各土层土壤有机质、全氮和碱解氮含量逐渐增加, 表层土壤养分增幅高于底层[40]。在采用围栏禁牧、布设沙障和补播草种方式进行沙化土壤的恢复过程中发现, 3年后植被恢复、土壤氮素得以积累, 土壤理化性质得到很大改善, 植被对土壤氮素含量有显著的影响[41]。在本研究中, 通过前期有机肥等养分向沙质土壤的施入和植被的建植, 各处理的植物界面对土壤界面中有机质、碱解氮、速效磷、速效钾含量普遍提高, 改良效应集中在0-20 cm土层, 沙质土壤养分状况得到改善, 其改良效果为Poa> Puc> Ely, 且3种草本植物改良效果间有显著性差异, 推测应该与不同植物的根系生长特性、根系在沙土中的分布及根系分泌物相关。
土壤酶主要来源于植物、微生物和土壤动物, 其中植物来源主要依靠植物根系向外分泌酶类, 土壤酶活性与地表植被、植被根际间具有一定的相关性[42, 43, 44]。土壤酶参与了土壤中所有的生化反应, 是土壤有机质分解和养分循环的主要生物学机制, 其活性反映了土壤中各种生物化学过程的方向和强度。土壤脲酶、转化酶和过氧化氢酶广泛存在于土壤中, 且是土壤中碳、氮、磷和钾素循环的关键酶。因此, 研究土壤中酶活性对探讨不同草本植物对沙土的改良作用程度具有重要意义。在本研究中, Ely、Puc、Poa共3个栽培草地处理均显著提高了土壤的酶活性, Poa处理效果最显著, 这说明随着生境的改善, 荒漠区土壤酶活性逐渐提高。主要原因是植被建植后, 土壤的理化性质包括土壤养分含量增加、养分有效性提高和土壤稳定性增强等, 这些为微生物的生存创造了适宜的条件, 从而有利于增加土壤酶的活性[45]。
在本研究中, 脲酶与土壤有机质、全氮、碱解氮、全磷、速效磷、全钾和速效钾显著正相关, 并集中体现在0-20 cm植物根系较密集的土层; 过氧化氢酶在Ely处理下在0-10 cm土层与土壤速效磷极显著正相关; 在Puc处理下在0-10 cm土层与速效磷和速效钾极显著正相关, 在10-20 cm土层与土壤全钾极显著负相关, 过氧化氢酶与其它养分之间复杂的关系, 可能是因为土壤中有机质的增加不仅会影响与之相关的过氧化氢酶活性, 而且还会矿化其它养分, 即一种养分的供应在调控另一种养分的矿化方面发挥一定的作用; 土壤转化酶对丰富土壤中能被植物和微生物利用的可溶性营养物质起着重要作用, 它与土壤中的全碳和全氮含量正相关, 是土壤中碳循环的重要指标。本研究中, 转化酶与土壤有机质呈现显著的正相关关系, 与其它营养成分之间无显著相关性, 可能与沙化土地环境(温度、湿度等)及沙化土壤恢复阶段相关, 有待进一步研究。
土壤养分与土壤酶活性之间存在非常密切而复杂的关系, 已有研究表明, 土壤酶活性与土壤养分含量之间均具有明显的平行性, 其活性大小往往与土壤营养呈正相关关系[46, 47, 48, 49]。在沙化治理过程中, 有机质往往在沙质土壤改良中起着重要作用, 有机质含量增加, 不仅增加了土壤中酶促底物的主要供源, 还可以作为土壤的有机载体, 保持土壤酶的活性和稳定性, 其转化分解过程会促使酶活性提高, 土壤酶同时也驱动着土壤中各种生物化学反应, 参与土壤中N、P、K等营养因素的循环。
不同草原植被下草甸土的土壤酶活性随着土壤微生物的增加不断增强, 且与有机质、全氮和全磷之间呈指数正相关关系。黑土和暗棕壤的土壤脲酶、磷酸酶、转化酶、过氧化氢酶活性与土壤有机质、全氮、全磷、碱解氮、速效磷含量的相关性均达显著水平[50]。土壤酶与土壤养分之间的这种显著相关性, 说明土壤酶作为土壤有机质分解和养分转化循环的催化剂, 与土壤养分含量和微生物生物量之间密切相关。
| [1] |
|
| [2] |
|
| [3] |
|
| [4] |
|
| [5] |
|
| [6] |
|
| [7] |
|
| [8] |
|
| [9] |
|
| [10] |
|
| [11] |
|
| [12] |
|
| [13] |
|
| [14] |
|
| [15] |
|
| [16] |
|
| [17] |
|
| [18] |
|
| [19] |
|
| [20] |
|
| [21] |
|
| [22] |
|
| [23] |
|
| [24] |
|
| [25] |
|
| [26] |
|
| [27] |
|
| [28] |
|
| [29] |
|
| [30] |
|
| [31] |
|
| [32] |
|
| [33] |
|
| [34] |
|
| [35] |
|
| [36] |
|
| [37] |
|
| [38] |
|
| [39] |
|
| [40] |
|
| [41] |
|
| [42] |
|
| [43] |
|
| [44] |
|
| [45] |
|
| [46] |
|
| [47] |
|
| [48] |
|
| [49] |
|
| [50] |
|
青海湖东沙化治理过程中土壤酶活性及养分含量特征
魏小星
Soil nutrient content and the activities of soil enzymes during the desertification restoration process to the east of Qinghai Lake
Wei Xiao-xing