


{kind=link}
{kind=link}
{kind=link}
{kind=link}
祁连山北坡3个季节性牧场夏季的土壤呼吸
[郭雅蓉1, 2 , 侯扶江1, 2  , 于应文
, 于应文1, 2 ]
, 于应文|
|


第一作者:郭雅蓉(1993-),女,山西长治人,在读硕士生,研究方向为作物-家畜综合系统。E-mail:[email protected]
以祁连山北坡高寒灌丛化草甸和高寒草原为对象,测定比较夏季、冬季和春秋季牧场的夏季土壤呼吸特征及与其影响因素之间的关系,为牧场碳汇管理提供依据。以高山灌丛化草甸为主的夏季牧场日均土壤呼吸速率(Rs)为0.26 μmol·(m2·s)-1,以高寒草原为主的冬季和春秋季牧场日均Rs分别为0.24和0.19 μmol·(m2·s)-1。随放牧增强,冬季牧场Rs减小,春秋季和夏季牧场Rs增加。冬季牧场Rs与土壤含水量(soil mosture,SM)显著正相关( P<0.05),Rs与放牧率(stocking rate,SR)和土壤温度(soil temperature,Ts)显著负相关( P<0.05);春秋季牧场Rs与SR、光合有效辐射(photosynthetically awailable radiation,PAR)、Ts显著正相关( P<0.05);春秋季牧场Rs与SM和群落盖度(community coverage,C)显著负相关( P<0.05)。结构模型方程表明,冬季牧场放牧率通过Ts对Rs的负向间接作用最强,SM对Rs正向的直接作用次之;春秋季牧场群落盖度对Rs负向直接作用最大,放牧率对Rs正向的直接作用次之。冬季牧场Rs可分别由 Rs=0 .436 -0 .012 Ts+0.018 SM( R2 =0 .911, P=0 .038)和 Rs=0.707+0.002 SR-0 .02 Ts( R2 =0 .775, P=0 .037) 估测;春秋季牧场可由 Rs=-0.239+0.003 PAR-0 .001 C( R2 =0 .979, P=0.004)估测。
To facilitate the management of carbon sink, soil respiration and factors affecting it were measured and analysed in summer pasture, winter pasture, and spring-autumn pasture on the northern slope of Qilian Mountains in summer. Daily soil respiration (Rs) was 0.24, 0.19, and 0.26 μmol·(m2·s)-1 in the winter pasture, spring-autumn pasture, and summer pasture, respectively. With increase in stocking rate, Rs decreased in winter pasture, whereas that in the spring-autumn pasture and summer pasture revealed the opposite trend. In winter pasture, Rs was significantly positively correlated with soil moisture (SM) and significantly negatively correlated with stocking rate (SR) and soil temperature (Ts) ( P<0.05). However, in spring-autumn pasture, Rs was significantly positively correlated with SR, photosynthetically available radiation (PAR), and SM ( P<0.05), but significantly negatively correlated with soil temperature (Ts) and community coverage (C) ( P<0.05). A structural model revealed that the indirect negative effect of SR by Ts on Rs was higher than the direct positive effect of SM in winter pasture. The direct negative effect of C on Rs was higher than the direct positive effect of SR in spring-autumn pasture. In winter pasture, Rs can be estimated better by Ts and SM, Rs=0.436-0.012 ST+0 .018 SM ( R2 =0 .775, P=0.037). In spring-autumn pasture, Rs can be estimated better by PAR and C, Rs=-0.239+0.003 PAR-0 .001 C ( R2 =0 .979, P=0.004).
土壤呼吸是其产生CO2的代谢过程, 是自养(根)和异养呼吸(土壤微生物)的总和[1]。陆地生态系统2/3的碳储存在土壤中[2], 土壤呼吸是一种重要的碳交换过程, 是土壤向大气输出碳的主要途径[3], 受土壤物理、化学和生物等因素的综合影响[4]。草地生态系统作为世界面积最大的陆地生态系统, 其土壤呼吸对调节全球碳循环具有重大作用[5], 对土壤有机质的矿化、异养代谢、土壤腐殖质和枯落物碳代谢、草地地下碳分配和生产力等有重要指示作用[6]。
土地利用方式显著影响草地生态系统土壤有机碳的贮存和释放[7], 而且是导致大气CO2浓度升高的重要原因之一[1]。放牧通过家畜采食、践踏和排泄粪便对土壤呼吸产生直接和间接作用[7]。家畜采食降低植被盖度和高度, 提高太阳透射率, 导致地表升温、蒸发增加, 促进土壤呼吸[8]。高强度践踏改变土壤紧实度、渗透率和透气性等物理结构以及生物土壤结皮、土壤微生物活性等, 导致土壤呼吸强度减弱[9, 10]。家畜排泄改善草地养分, 影响土壤微生物活性和根系代谢, 从而加快生态物质循环和能量流动, 进而促进土壤呼吸[11]。适度放牧促进草地生物量、根系和凋落物数量、土壤动物和微生物多样性的提高, 从而增强土壤呼吸; 不合理的放牧导致草地植被稀疏, 草地生物量下降, 从而降低土壤呼吸速率[12, 13]。青藏高原高山草甸的放牧强度降低约50%, 土壤释放的CO2大约翻一翻[14]。新疆天山高寒草原短期禁牧, 土壤CO2和N2O排放与长期禁牧和自由放牧差异不显著[15]。
国内外放牧强度对土壤呼吸的影响研究很多[2, 3, 16, 17, 18, 19, 20, 21, 22, 23, 24, 25, 26, 27, 28, 29], 但在牧场尺度上, 放牧季节对草原土壤呼吸的影响报道较少, 且放牧强度多以定性的过牧或重牧、中牧、轻牧、禁牧或围封等为主, 而放牧强度与土壤呼吸之间的定量关系尚不明确。国内对于高寒草甸研究较多[15, 23, 29, 30, 31], 但对高寒灌丛化研究较少。为此, 研究祁连山北坡不同季节牧场土壤呼吸特征及与影响土壤呼吸因素之间的相互关系, 并建立预测模型, 为牧场碳汇管理提供理论依据, 还可为牧场科学管理决策提供理论支持。
研究区位于甘肃省肃南裕固族自治县甘肃马鹿(Cervus elaphus kansuensis)养殖场, 有3个季节牧场, 地处祁连山中段北麓, 地理坐标38.8° N、99.6° E附近, 平均海拔2 850 m。年均温3.6 ℃, 年均降水量253.0 mm, 主要集中在6月-9月。牧草一般4月下旬返青, 7月上旬进入生长旺盛期, 9月上旬开始枯黄[19]。
试验设在甘肃马鹿的冬季牧场(winter pasture, WP)、春秋季牧场(spring-autumn pasture, SAP)和夏季牧场(summer pasture, SP)。根据草原综合顺序分类法[32], 冬季和春秋季牧场属于寒温微干山地草原类, 土壤为山地栗钙土[33]; 夏季牧场属于亚高山灌丛草甸, 土壤属于高山灌丛草甸土。冬季和春秋季牧场主要植物种为紫花针茅(Stipa purpurea)、短花针茅(S. breviflora)、扁穗冰草(Agropyron cristatum)、醉马草(Achnatherum inebrians)、冷蒿(Artemisia frigida)、赖草(Leymus secalinus)、银灰旋花(Convolvulus ammannii)和其它一些植物[9, 34]; 夏季牧场主要植物种为鬼箭锦鸡儿(Caragana jubata); 冬季牧场的醉马草、甘肃马先蒿(Pedicularis kansuensis)、狼毒(Stellera chamaejasme)和银灰旋花等毒杂草分布广泛, 在牧场入口附近甚至是优势种或次优势种, 退化严重。醉马草仅分布在春秋季牧场入口附近, 纵深区域较少, 退化较轻; 夏季牧场畜圈附近退化严重, 醉马草为优势种[35]。
冬季和春秋季牧场沿马鹿采食的主要路线在距牧场入口0、300、600、900、1 200和1 500 m处共设6个样地, 向外呈放射状形成放牧率由高到低的牧压梯度。1个家畜单位(animal unit, AU)为1头成年甘肃马鹿母鹿(活体重200 kg左右)带一只哺乳期的小鹿。依据家畜体量或采食量, 将各年龄段马鹿换算为标准家畜单位[34, 36, 37]。放牧率以单位放牧地面积上家畜单位与放牧时间(月month, M)的乘积(AUM)表示, 分别得到冬季和春秋季牧场的放牧率(表1)[34, 36, 37]。1999年夏季从放牧开始, 直至放牧结束, 每隔2 h观察一次马鹿的放牧行为, 记录样地内马鹿数量和年龄。
冬季和春秋季牧场各样地放牧率(stocking rate, SR)计算公式:
SRi=SR×
式中:SRi为样地i的放牧率, SR为整个牧场的放牧率, Fi为家畜在样地i的出现频率, n为样地的总数[36, 37]。
夏季牧场由南北走向的两个小山及其间谷地组成, 距离畜圈50和600 m, 分别在东西方向上设置两条样带, 600 m样带位于50 m样带南侧, 海拔较50 m样带高50 m左右。每条样带分为坡顶、坡中、坡谷; 根据坡向分为阳坡和阴坡(表2)。
应用CI-301PS闭路系统的CI-301SR土壤呼吸室测定土壤呼吸。冬季牧场, 1999年7月06:00-21:00, 每隔2 h测定一次, 每样地每次随机选择5个样点, 每样点测定5次, 取平均值。春秋季牧场和夏季牧场, 09:00-11:00测定一次, 每样地随机布设5个样点, 每样点测5个数据。同时测定气温、光照、土壤地表温度、土壤含水量等数据。
土壤呼吸速率计算公式:
Rs=12 323.35× V× P/Δ t× Ta× A .
式中:Rs为土壤呼吸速率[μ mol· (m2· s)-1], V为体积流速(L· min-1), P为大气压力(Pa), Δ t为测定间隔时间(Δ t=30 s), Ta为空气湿度(K), A为呼吸室面积(A=74.5 cm2)。
冬季牧场日均土壤呼吸速率:用测定的土壤呼吸速率对时间积分, 取其时间平均值。
冬季和春季牧场在每个样地随机取5个(1 m× 1 m)样方, 统计每个样方中的植物种类, 记录每个植物种的投影盖度, 总盖度=各个植物种的分盖度之和。
| 表1 祁连山北坡不同放牧率下冬季和春秋季牧场植物群落 Table 1 Plant community in winter and spring-autumn pasture under different stocking rates on the northern slope of Qilian Mountains |
| 表2 祁连山北坡夏季牧场不同坡位植物群落 Table 2 Plant community on the different slope positions in summer pasture on the northern slope of Qilian Mountains |
土样采集与群落盖度同步, 每样地用直径5 cm土钻取15钻, 0-40 cm深度每10 cm为一层, 筛出根系, 测定根系生物量[38]。
用SPSS 20.0软件, 对祁连山北坡夏季不同季节牧场影响土壤呼吸因子及其与土壤呼吸之间进行相关性分析; 运用LSD法分别对夏季牧场相同距离不同坡位、不同距离相同坡位土壤呼吸进行多重比较; 利用通径分析方法, 分析影响土壤呼吸因子分别在冬季和春秋季牧场对土壤呼吸的直接和间接作用; 采用多元回归方法, 建立影响土壤呼吸因子与土壤呼吸之间的回归模型。用Microsoft Excel 2010软件绘图。
冬季牧场300 m样地土壤呼吸日动态具有代表性, 基本呈单峰曲线变化, 14:00左右最高(图1), 与土壤温度变化一致。
 | 图1 冬季牧场土壤呼吸和土壤温度日动态Fig. 1 Daily dynamics of soil respiration and soil temperature in winter pasture on the northern slope of Qilian Mountains |
随放牧率增加, 冬季牧场日均土壤呼吸速率下降, 春秋季牧场呈增加变化(图2)。放牧率增加1 AUM· hm-2, 冬季和春秋季牧场土壤呼吸分别减少0.014 2 μ mol· (m2· s)-1和增加0.025 7 μ mol· (m2· s)-1。
 | 图2 冬季牧场和春季牧场放牧率与土壤呼吸速率的关系Fig. 2 The relationship between stocking rate and soil respiration in winter and spring-autumn pasture on the northern slope of Qilian Mountains |
夏季牧场, 相同坡位, 离畜圈50 m的样地土壤呼吸速率高于离畜圈600 m的样地(P< 0.05)(表3); 相同距离不同坡位土壤呼吸不同。夏季牧场阴坡坡顶日均土壤呼吸速率[Rs=0.318 μ mol· (m2· s)-1 ]> 坡谷[Rs=0.214 μ mol· (m2· s)-1 ]> 坡中[Rs=0.165 μ mol· (m2· s)-1 ], 阳坡坡中日均土壤呼吸速率[Rs=0.308 μ mol· (m2· s)-1 ]> 坡顶[Rs=0.243 μ mol· (m2· s)-1 ]> 坡谷[Rs=0.214 μ mol· (m2· s)-1 ]。
| 表3 祁连山北坡夏季牧场不同坡位土壤呼吸速率 Table 3 Soil respiration on the different slope positions in summer pasture on the northern slope of Qilian Mountains |
冬季牧场土壤呼吸速率与放牧率、土壤含水量和土壤温度显著相关(P< 0.05)(表4), 放牧率与土壤0-10 cm温度显著负相关(P< 0.05); 冬季牧场土壤呼吸速率与其含水量显著正相关(P< 0.05), 与放牧率和土壤温度显著正相关(P< 0.05)。
| 表4 祁连山北坡冬季和春秋季牧场土壤呼吸影响因素之间的相关性分析 Table 4 Correlation among the factors of affecting soil respiration in winter and spring-autumn pasture on the northern slope of Qilian Mountains |
影响春秋季牧场土壤呼吸的因素较多, 放牧率、光合有效辐射和土壤温度与土壤呼吸显著正相关(P< 0.05), 土壤含水量、群落盖度与土壤呼吸显著负相关(P< 0.05)。
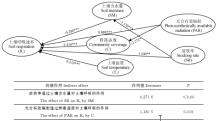
冬季牧场土壤呼吸通径分析显示(图3), 土壤含水量和温度对冬季牧场土壤呼吸是直接作用, 放牧对土壤呼吸是间接作用, 土壤温度相较于土壤含水量对土壤呼吸的直接作用较大, 说明冬季牧场日均土壤呼吸速率土壤0-10 cm温度变化较敏感。放牧主要通过改变土壤0-10 cm温度间接影响冬季牧场土壤呼吸。
 | 图3 祁连山北坡冬季牧场土壤呼吸通径分析注:实线为直接影响。* , * * , * * * 分别表示在0.05、0.01和0.001水平上显著。下图同。Fig. 3 The path analysis of soil respiration in winter pasture on the northern slope of Qilian MountainsNote: Solid lines indicate direct effects. * , * * , and * * * indicate significant relationship at 0.05, 0.01, and 0.001 levels, respectively. similarly for the Fig. 4. |
通径分析显示, 春秋季牧场放牧率、光合有效辐射、土壤温度、土壤含水量和群落盖度均对土壤呼吸产生直接作用, 放牧率和光合有效辐射对土壤呼吸有间接作用(图4)。群落盖度对春秋季牧场的土壤呼吸的直接作用最大, 放牧次之; 光合有效辐射通过群落盖度对土壤呼吸的间接作用较大, 放牧通过土壤含水量对土壤呼吸的间接作用极显著。可见, 群落盖度是春秋季牧场土壤呼吸主要限制因子。
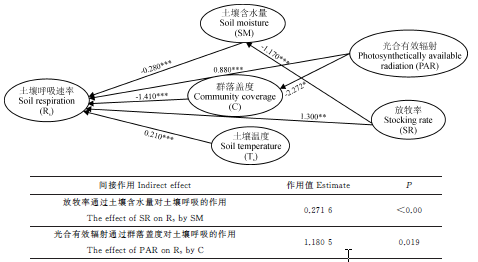 | 图4 祁连山北坡春秋季牧场土壤呼吸通径分析Fig. 4 The path analysis of soil respiration in spring-autumn pasture on north slope of Qilian Mountains |
冬季牧场分别以放牧率和土壤温度、土壤温度和土壤含水量预测土壤呼吸的两个决定系数均在0.05水平上显著, 说明冬季牧场土壤呼吸可由其与放牧率和土壤温度、土壤温度和土壤含水量之间的多元回归模型很好地预测(表5)。春秋季牧场光合有效辐射和群落盖度预测土壤呼吸的决定系数在0.01水平上显著, 说明春秋季牧场土壤呼吸可由其与光合有效辐射和群落盖度之间的多元回归模型很好地预测。
| 表5 祁连山北坡土壤呼吸多元预测模型 Table 5 The estimated model of soil respiration on north slope of Qilian Mountains |
冬季和春秋季牧场在夏季的土壤呼吸速率与放牧率变化趋势相反, 可能由于冬季牧场放牧较重, 破坏了土壤理化性状, 一方面土壤水分和有机碳下降, 抑制土壤微生物和根系活动[37, 38], 另一方面牧草地上部分光合物质同化下降, 向根系物质运输减少, 使根系活动减弱[33, 34]; 春秋季牧场放牧较轻, 家畜春季适度放牧减少了群落盖度, 改善了群落微气候, 到达地面的太阳辐射增强, 土壤升温较快, 土壤呼吸强度随之增大[39]。春秋季牧场土壤呼吸速率低于冬季牧场, 表明其碳汇功能较强, 可能与放牧强度较低有关。
从土壤呼吸速率、土壤含水量和土壤温度相关性来看, 冬季牧场土壤呼吸速率与土壤温度显著负相关, 与短花针茅荒漠草原不同放牧制度下土壤呼吸速率与土壤温度相关性[28]相同, 而亚高山草甸在生长季节的土壤呼吸速率与土壤温度正相关[29], 这可能是由于土壤呼吸受多种生物、非生物因素影响有关。一些研究表明, 当土壤湿度成为冬季牧场限制因子时, 土壤呼吸对温度的敏感性会降低[31, 40]; 当土壤温度较低时, 增加冠层盖度可提高土壤湿度, 从而促进土壤呼吸[1]。虽然冬季和春秋季牧场的土壤含水量与土壤温度均呈负相关关系, 但冬季牧场的相关性不显著, 而春秋季牧场呈显著负相关关系; 这可能是土壤呼吸在小尺度范围内受地形地貌[40]、植被[41]、土壤质地[42]以及土地利用方式[43]的影响, 从而土壤呼吸表现出明显空间变化特征; 另一方面, 由于不同植物、群落、土壤微气候存在一定差异, 而土壤温度和水分均可通过影响土壤中的微生物活性及植被根系生长直接影响土壤呼吸作用[44, 45]。
在内蒙古短花针茅荒漠草原, 放牧削弱了土壤水分对土壤呼吸速率的影响, 可能是放牧样地由于牲畜的采食和踩踏, 地上生物量和凋落物等减少, 导致土壤表层温度易升高, 土壤水分降低[46]。但本研究中, 冬季牧场放牧促进土壤表层温度对土壤呼吸速率的影响, 从而对土壤呼吸产生影响。
夏季牧场阴坡坡顶日均土壤呼吸速率大于坡谷大于坡中, 阳坡坡顶日均土壤呼吸速率小于坡中但大于坡谷, 这可能因为高寒灌丛草甸土壤水分充足, 热量是土壤呼吸的限制因子, 放牧强度越大, 群落透光性越好, 土壤升温越快, 因此太阳辐射量越大, 日均土壤呼吸速率越大[6, 47]。
随放牧率增大, 冬季牧场土壤呼吸速率减小, 春秋季和夏季牧场土壤呼吸速率增大。土壤呼吸主要影响因子, 冬季牧场为放牧率、土壤含水量与土壤温度, 春秋季牧场为放牧率、光合有效辐射、土壤温度、土壤含水量和群落盖度, 夏季牧场为坡位。冬季牧场的土壤含水量和土壤温度分别对其土壤呼吸起正向直接作用和负向直接作用, 放牧率通过土壤表层温度对土壤呼吸起负向间接作用; 春秋季牧场放牧率、土壤温度、土壤含水量、群落盖度主要对土壤呼吸起直接作用, 光合有效辐射对土壤呼吸主要起间接作用; 夏季牧场的土壤呼吸速率阴坡坡顶> 坡谷> 坡中, 阳坡为坡中> 坡顶> 坡谷。冬季牧场Rs可分别由Rs=0.436-0.012Ts+0.018SM和Rs=0.707+0.002SR-0.02Ts估测; 春秋季牧场Rs可由Rs=-0.239+0.003PAR-0.001C估测。
(责任编辑 武艳培)
The authors have declared that no competing interests exist.
| [1] |
|
| [2] |
|
| [3] |
|
| [4] |
|
| [5] |
|
| [6] |
|
| [7] |
|
| [8] |
|
| [9] |
|
| [10] |
|
| [11] |
|
| [12] |
|
| [13] |
|
| [14] |
|
| [15] |
|
| [16] |
|
| [17] |
|
| [18] |
|
| [19] |
|
| [20] |
|
| [21] |
|
| [22] |
|
| [23] |
|
| [24] |
|
| [25] |
|
| [26] |
|
| [27] |
|
| [28] |
|
| [29] |
|
| [30] |
|
| [31] |
|
| [32] |
|
| [33] |
|
| [34] |
|
| [35] |
|
| [36] |
|
| [37] |
|
| [38] |
|
| [39] |
|
| [40] |
|
| [41] |
|
| [42] |
|
| [43] |
|
| [44] |
|
| [45] |
|
| [46] |
|
| [47] |
|
祁连山北坡3个季节性牧场夏季的土壤呼吸
郭雅蓉, 于应文
Soil respiration in three types of seasonal pastures in summer on the northern slope of Qilian Mountains
Guo Ya-rong, Yu Ying-Wen