




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
盐胁迫对红豆草幼苗生长和离子积累及分配的影响
[伍国强1  , 贾姝
, 贾姝1 , 刘海龙1 , 王春梅2 , 李善家1 ]
, 贾姝|
|


以3周龄红豆草( Onobrychis viciaefolia)幼苗为材料,研究了不同浓度(0、5、25、50、100和200 mmol·L-1)NaCl及50 mmol·L-1处理不同时间(0、1、3、5、7和9 d)对其生长和离子积累及分配的影响,以期解析红豆草响应盐胁迫的生理机制。结果表明,与对照(0 mmol·L-1)相比,5、25和50 mmol·L-1 NaCl对红豆草幼苗生长影响不大;而100和200 mmol·L-1明显抑制其生长。随着盐浓度增加,红豆草地上部和根Na+浓度呈显著增加趋势( P<0.05),其中50200 mmol·L-1下,地上部Na+浓度较对照增加1439倍。特别是200 mmol·L-1使根Na+净吸收速率较对照增加了35倍,使整株总Na+量和地上部Na+相对分配比例分别增加了93%和45%;而显著降低了根的K+、Na+选择性运输能力。在50 mmol·L-1 NaCl下,随着处理时间延长,地上部和根Na+浓度也呈逐渐增加趋势,但根中的Na+到7 d时达到最大,然后呈下降趋势;相反,K+浓度均呈缓慢下降趋势,且根中的浓度始终高于地上部。由此可见,在550 mmol·L-1NaCl下,红豆草通过维持其体内K+、Na+稳态平衡抵御盐胁迫;而在100和200 mmol·L-1下,红豆草地上部积累大量Na+,但其叶片Na+区域化及K+选择性转运能力较弱,导致植株体内Na+、K+稳态紊乱,产生离子毒害,从而抑制其生长。
In this study, we investigated the effects of different concentrations of NaCl (0, 5, 25, 50, 100 and 200 mmol·L-1) over different time periods (0, 1, 3, 5, 7 and 9 d), after 50 mmol·L-1 NaCl treatment, on growth, ion accumulation, and distribution in 3-week-old sainfoin ( Onobrychis viciifolia) seedlings. Results showed that concentrations of 5, 25, and 50 mmol·L-1 did not significantly affect plant growth, while concentrations of 100 and 200 mmol·L-1 significantly inhibited growth when compared with that in the control (0 mmol·L-1)( P<0.05). With the increase of NaCl concentrations, Na+ concentrations in both shoot and root tended to increase. Compared with the control, the addition of 50200 mmol·L-1 NaCl increased shoot Na+ concentration by 1439 times, respectively. In particular, concentrations of 200 mmol·L-1 significantly increased root Na+ net uptake rates, total Na+ amount, and relative shoot Na+ distribution by 35-fold, 93% and 45%, respectively; whereas it reduced selective transport capacity for K+ over Na+ from roots to shoots. At 50 mmol·L-1 NaCl, with increased treatment time, Na+ accumulation in both shoot and root tended to increase, but that in the roots reached a peak on the 7th day and reduced subsequently. In contrast, K+ concentrations showed a decreasing trend, to a larger degree in root than in shoot. These results indicate that under NaCl concentrations of 550 mmol·L-1, sainfoin plants could adapt to salt stress by maintaining K+ and Na+ homeostasis; while under 100 and 200 mmol·L-1, plants passively accumulated a large amount of Na+ in shoots, where ions compartmentation and selective transport capacity for K+ over Na+ were weak, thus disrupting the balance of Na+ and K+ and resulting in osmotic stress and growth inhibition.
盐胁迫是影响植物生长和作物产量的主要非生物因素之一[1, 2, 3], 全球约有20%的灌溉地遭受盐渍化的威胁[4]。我国盐碱地面积约为3.5× 107 h
红豆草(Onobrychis viciaefolia)是一种优良的多年生豆科牧草, 被誉为“ 牧草皇后” [14, 15], 在我国北方干旱、半干旱地区广泛种植。其根系发达, 根上有块状根瘤, 可以生物固氮, 有效提高土壤肥力; 营养丰富, 产量高, 含有大量单宁, 牲畜喜食且不引起膨胀病; 花色粉红艳丽, 气味芳香, 也是一种较为理想的绿化、美化和观赏植物[16, 17, 18, 19]。目前, 有关盐胁迫对红豆草种子萌发、生活力和其固氮作用的研究报道较多[20, 21, 22, 23, 24]。包桂荣等[20]采用NaCl和Na2CO3各6个浓度梯度(0、500、1 000、2 000、3 000和4 000 mg· L-1)溶液对红豆草种子进行处理, 发现低盐浓度(500 mg· L-1)能提高种子发芽率, 但高盐浓度则降低种子发芽率和发芽势及胚根、胚芽长度。尽管有关红豆草耐盐机制方面的研究取得了一些进展[16], 然而盐胁迫下其体内离子积累及分配规律尚不清楚。鉴于此, 本研究以3周龄红豆草幼苗为材料, 探究不同浓度NaCl处理下其生长和不同组织Na+、K+的积累和分配, 以期进一步揭示红豆草响应盐胁迫的生理机制。
供试红豆草品种为“ 甘肃” , 种子由兰州大学草地农业科技学院李彦忠教授馈赠。试验于2015年3月-9月在兰州理工大学生命科学与工程学院植物培养室进行。挑选籽粒饱满、均匀一致的种子, 播种在装有蛭石的塑料盘(5 cm× 5 cm× 5 cm, 每盘8孔)中, 每孔播56粒种子, 浇灌蒸馏水进行萌发, 待两片子叶露出蛭石后, 浇灌1/2 Hoagland营养液进行培养。营养液配方为:2 mmol· L-1 KNO3, 0.5 mmol· L-1 NH4H2PO4, 0.25 mmol· L-1 MgSO4· 7H2O, 0.1 mmol· L-1 Ca(NO3)2· 4H2O, 0.5 mmol· L-1 Fe-citrate, 92 μ mol· L-1 H3BO3, 18 μ mol· L-1 MnCl2· 4H2O, 1.6 μ mol· L-1 ZnSO4· 7H2O, 0.6 μ mol· L-1 CuSO4· 5H2O, 0.7 μ mol· L-1 (NH4)6M· 7O24· 4H2O。待第1片真叶出现后, 间苗, 每孔留3株。培养室昼夜温度 (28± 2) ℃/(23± 2) ℃, 光照时间16 h· d-1, 光强为500600 μ mol· (m2· s)-1, 空气相对湿度为60%80%。每两天更换一次营养液。
3周龄红豆草幼苗按如下方案处理:1)分别用含0(对照)、5、25、50、100和200 mmol· L-1 NaCl的1/2 Hoagland营养液处理7 d; 2)用含50 mmol· L-1 NaCl的1/2 Hoagland营养液分别处理0(对照)、1、3、5、7和9 d。每个处理设8次重复。每两天更换一次处理液, 以保持恒定的NaCl浓度。
鲜重和干重:参考Wu等[25]的方法略有修正, 将植物材料用蒸馏水冲洗表面灰尘和残留盐分后, 用吸水纸吸干其表面水分, 将其分为地上部和根, 分别称取鲜重; 然后将样品装在信封中, 杀青后, 置于80 ℃烘干至恒重, 称取干重。
Na+、K+浓度和K+/Na+:参考Yue等[26]的方法并略有修正, 将恒重的干样品在研钵中, 研磨至粉末状后, 放入试管中, 添加 100 mmol· L-1冰乙酸, 密封试管, 置于96 ℃沸水中水浴2 h。过滤后, 上清液稀释适当倍数后, 在火焰光度计(2655-00, Cole-Parmer Instrument Co., USA)上测定Na+和K+浓度。K+/Na+计算方法参考文献[27]。K+、Na+选择性吸收(SA)和选择性运输(ST)能力及根K+、Na+净吸收速率的计算方法参考文献[27-28]。
采用SPSS 19.0软件(SPSS Inc., USA)对试验数据进行单因素方差分析, 显著性差异分析采用Duncan检验进行(P< 0.05), 采用Excel 2007制图。
与对照(0 mmol· L-1)相比, 5、25和50 mmol· L-1 NaCl对红豆草地上部鲜重没有显著性影响(P> 0.05); 在100和200 mmol· L-1 NaCl下, 植株地上部鲜重较对照分别降低了21%和23%(P< 0.05)(图1)。50、100和200 mmol· L-1NaCl下, 幼苗地上部干重较对照分别显著下降了29%、35%和28%(P< 0.05)(图1)。然而, 盐浓度的变化对红豆草幼苗根的鲜重和干重无显著影响(P> 0.05)。
 | 图1 不同浓度NaCl对红豆草鲜重和干重的影响 注:不同小写字母表示处理间差异显著(P< 0.05)。下同。Fig. 1 Effect of different concentrations of NaCl on fresh and dry weight in sainfoin Note: Different lowercase letters indicate significant difference at the 0.05 level; similarly for the following figures. |
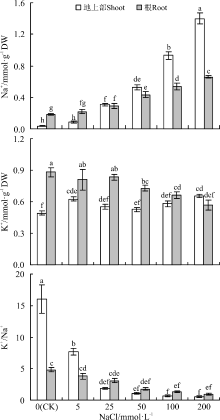
随着NaCl浓度的增加, 无论地上部还是根中的Na+浓度均呈显著增加趋势(P< 0.05), 但相比之下, 地上部增加的幅度则更大(图2)。50、100和200 mmol· L-1 NaCl使地上部Na+浓度较对照分别增加了14、26和39倍, 而使根中K+浓度分别显著降低了17%、25%和36%。与对照相比, 不同浓度NaCl处理均显著降低了幼苗地上部和根K+/Na+。
 | 图2 不同浓度NaCl对红豆草Na+、K+浓度和K+/Na+的影响Fig. 2 Effect of different concentrations of NaCl on Na+, K+ concentrations and K+/Na+ in sainfoin |
进一步分析发现, 随着NaCl浓度的增加, 红豆草植株K+、Na+选择性运输能力呈下降趋势(图3), 而K+、Na+选择性吸收能力和Na+净吸收速率均呈逐渐增加趋势(P< 0.05)(图3)。尤其是在200 mmol· L-1 NaCl下, 根Na+净吸收速率较对照增加了35倍, 而K+、Na+选择性转运能力下降了82%。
在200 mmol· L-1 NaCl处理下, 植株总Na+含量和地上部Na+相对分配比例较对照分别增加了93%和45%, 而根Na+相对分配比例降低了77%(P< 0.05)(表1)。50、100和200 mmol· L-1 NaCl处理使总K+含量较对照显著降低, 但地上部的K+相对分配比例显著较对照增加, 而根中的分配比例仅200 mmol· L-1 NaCl处理较对照则显著下降。由此可见, 盐处理下红豆草一方面通过根吸收大量的Na+转运至地上部, 另一方面在一定程度上维持其体内K+的稳态平衡。
 | 图3 不同浓度NaCl对红豆草选择性吸收(SA)、选择性运输(ST)值和Na+净吸收速率的影响Fig. 3 Effect of different concentrations of NaCl on selective absorption(SA), selective transportation(ST), and Na+ net uptake rate in sainfoin |
| 表1 不同浓度NaCl对红豆草Na+、K+相对分配比例的影响 Table 1 Effect of different concentrations of NaCl on Na+, K+ relative distribution in sainfoin |
在50 mmol· L-1 NaCl下, 处理3 d时植株地上部鲜重和干重较对照(0 d)分别增加了27%和77%(P< 0.05)(图4)。处理3 d时根鲜重达到最大值, 较对照增加了49%(P< 0.05)(图4)。
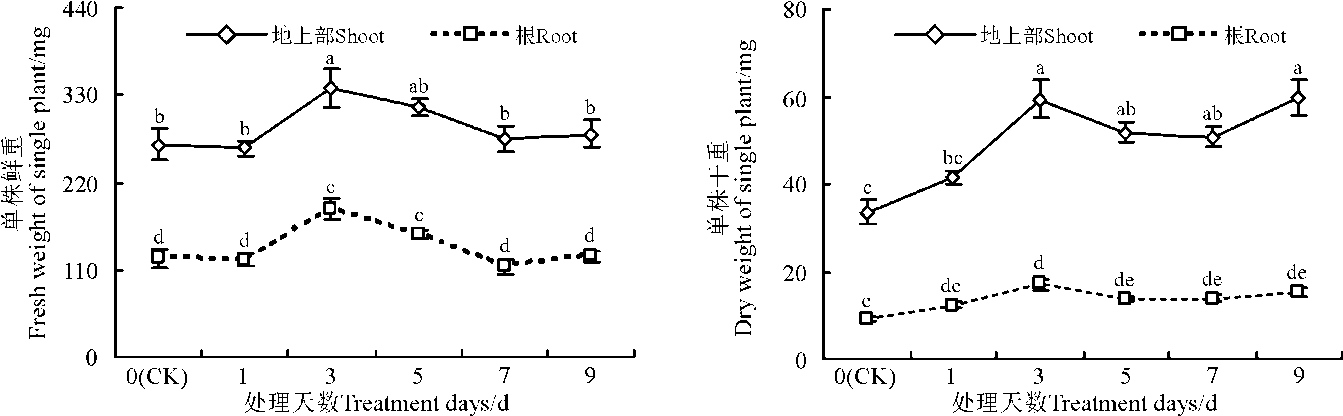 | 图4 50 mmol· L-1 NaCl处理不同时间对红豆草鲜重和干重的影响Fig. 4 Effect of different lasting times on fresh and dry weight in sainfoin under 50 mmol· L-1 NaCl |
在50 mmol· L-1 NaCl下, 随着处理时间延长, 地上部和根Na+浓度均呈逐渐增加趋势, 而根中Na+浓度到处理7 d时达到峰值, 随后呈下降趋势(图5)。在处理5 d之前根Na+浓度显著高于地上部(P< 0.05), 7 d时二者无显著差异(P> 0.05), 而9 d时地上部显著高于根(图5)。另外, 地上部和根K+浓度随着处理时间的延长呈缓慢下降趋势; 相比之下, 根中的K+浓度始终高于地上部(图5)。处理1 d时, 地上部和根K+/Na+较对照显著降低, 但地上部降低幅度更大; 随后呈缓慢下降趋势, 二者维持在同一水平(图5)。
 | 图5 50 mmol· L-1 NaCl处理不同时间对红豆草Na+ 、K+ 和K+/Na+的影响Fig. 5 Effect of different lasting times on Na+, K+, and K+/Na+ in sainfoin under 50 mmol· L-1 NaCl |
在50 mmol· L-1 NaCl下, 随着处理时间的延长, 植株K+、Na+选择性吸收和运输能力均呈显著下降趋势(P< 0.05), 处理9 d时其较对照分别降低了82%和76%(图6)。根Na+净吸收速率在处理5 d时达到最小值, 与处理1 d时相比下降了14%(图6)。从处理1 d开始, 根K+净吸收速率呈逐渐降低趋势, 处理9 d时较对照降低了91%(P< 0.05)(图6)。
 | 图6 50 mmol· L-1 NaCl处理不同时间对红豆草SA、ST、Na+和K+净吸收速率的影响Fig. 6 Effect of different lasting times on SA, ST, Na+ and K+ net uptake in sainfoin under 50 mmol· L-1 NaCl |
在50 mmol· L-1 NaCl下, 处理9 d时植株总Na+含量和地上部Na+相对分配比例较对照分别增加了95%和35%, 而根中相对分配比例降低了66%(P< 0.05)(表2)。9 d时植株总K+含量较对照增加15%(表2)。由此可见, 随着处理时间长, 植株体内Na+的积累量随之增加, 并将其大多数积累在地上部。
| 表2 50 mmol· L-1 NaCl处理不同时间对红豆草Na+、K+相对分配比例的影响 Table 2 Effects of different lasting times on Na+, K+relative distribution in sainfoin under 50 mmol· L-1 NaCl |
红豆草是一种优质的豆科牧草, 主要种植在内蒙古、新疆、陕西、甘肃和青海等省(区)[19], 这些地区由于频繁性季节交替作用, 其环境变化常引发土壤干旱和盐碱化, 使红豆草种子萌发和幼苗生长受到严重威胁。生物量是植物对盐胁迫反应的综合体现, 也是植物耐盐性的重要指标[29]。对于大多数植物而言, 当外界Na+浓度超过一定水平时会产生毒害作用。然而, 对一些盐生植物来说, Na+又是一种有益离子[3, 25]。在本研究中, 当盐浓度低于50 mmol· L-1时, 红豆草生长良好, 相似结果在霸王(Zygophyllum xanthoxylum)、细叶滨藜(Atriplex gmelini)和盐地碱蓬(Suaeda salsa)中均有报道[26, 30, 31]。盐胁迫下, 这些植物一方面可通过增加叶片的肉质化程度、提高组织含水量; 另一方面促进植株新叶的生长、提高其生物量, 将叶片中的 Na+稀释到毒害水平以下, 以适应盐渍生境[26, 30, 31]。与对照相比, 添加5、25和50 mmol· L-1 NaCl对红豆草地上部鲜重没有显著影响, 而100和200 mmol· L-1分别显著降低了21%和23%(P< 0.05)(图1), 50~200 mmol· L-1显著降低了幼苗地上部干重(图1)。这些结果表明, 红豆草对盐胁迫的耐受能力有限, 浓度高于50 mmol· L-1 的盐溶液则抑制其生长。
盐胁迫下, 植物体内积累的大量Na+会抑制细胞对K+的吸收, 从而使K+含量下降, 导致K+、Na+稳态失衡[32, 33]。赵昕等[34]研究发现, 盐胁迫(100和200 mmol· L-1)下盐生植物盐芥(Thellungiella halophila)从介质中吸收的大多数Na+积累在根中, 限制其转运到叶中, 使叶中保持较高K+/Na+, 从而适应盐渍环境。叶武威等[35]报道认为, 棉花(Gossypium hirsutum)具有较强的离子区域化能力, 使Na+积累在在茎和叶柄中, 而K+转运至叶片中, 以维持光合器官较高K+/Na+。此外, 霸王、盐地碱蓬、甜菜(Beta vulgaris)和枸杞(Lycium barbarumin)等能从介质中吸收大量Na+, 并将其区域化至叶片的液泡中, 一方面可降低细胞渗透势, 增强细胞吸水能力; 另一方面可降低Na+在细胞质中过量积累, 减轻对细胞的毒害作用[26, 31, 36, 37]。本研究发现, 当盐浓度低于50 mmol· L-1时, 幼苗根中的Na+积累量高于地上部, 由此说明红豆草能够限制的Na+从根向地上部转运, 从而使叶片中维持较低的Na+水平。然而, 当外界NaCl浓度为100和200 mmol· L-1或者50 mmol· L-1处理9 d时, 幼苗地上部的Na+含量显著高于根部, 由此说明, 当红豆草被动吸收的Na+超过根容纳范围时, 只能将其转运至地上部叶中, 但红豆草液泡区域化能力有限, Na+便会在细胞质积累, 对植株造成离子毒害, 从而抑制其生长。
K+、Na+选择性吸收和选择性转运能力是衡量植物耐盐性的重要指标, 不同植物对离子的选择性吸收和转运能力有所不同[38]。在本研究中, 在200 mmol· L-1下, K+、Na+选择性吸收能力最大, 类似结果在沙芥(Pugionium cornutum)中也有报道[39]。然而, 与50 mmol· L-1处理相比, 在200 mmol· L-1下红豆草地上部K+显著增加24%, 而K+/Na+无显著差异(图2), 表明高浓度NaCl下红豆草在一定程度上能通过限制Na+、增强K+的选择性吸收, 使叶中积累更多的K+, 维持恒定的K+/Na+。Wu等[40]研究表明, 5~200 mmol· L-1 NaCl处理下, 向日葵(Helianthus annuus)根部K+的选择性吸收能力显著增强, 而对K+的选择性转运能力显著降低。在本研究中, 不同盐浓度处理下, ST均显著性下降, 表明红豆草根系限制Na+、促进K+向地上部运输的能力较弱。
综上所述, 当盐浓度低于50 mmol· L-1时, 红豆草将吸收的Na+积累在根中, 限制Na+转运至地上部, 以保护叶片免受Na+毒害, 从而维持其正常生长。然而, 当盐浓度为100和200 mmol· L-1时, 红豆草根部被动吸收的大量Na+转运并积累在地上部叶片, 但其叶片Na+区域化及对K+的选择性转运能力较弱, 使植株体内K+、Na+稳态失衡, 产生离子毒害, 从而抑制其生长。
(责任编辑 武艳培)
The authors have declared that no competing interests exist.
| [1] |
|
| [2] |
|
| [3] |
|
| [4] |
|
| [5] |
|
| [6] |
|
| [7] |
|
| [8] |
|
| [9] |
|
| [10] |
|
| [11] |
|
| [12] |
|
| [13] |
|
| [14] |
|
| [15] |
|
| [16] |
|
| [17] |
|
| [18] |
|
| [19] |
|
| [20] |
|
| [21] |
|
| [22] |
|
| [23] |
|
| [24] |
|
| [25] |
|
| [26] |
|
| [27] |
|
| [28] |
|
| [29] |
|
| [30] |
|
| [31] |
|
| [32] |
|
| [33] |
|
| [34] |
|
| [35] |
|
| [36] |
|
| [37] |
|
| [38] |
|
| [39] |
|
| [40] |
|
盐胁迫对红豆草幼苗生长和离子积累及分配的影响
伍国强, 贾姝
Effect of salt stress on growth, ion accumulation, and distribution in sainfoins seedlings
Wu Guo-qiang, Jia Shu