
{kind=link}
两种形态氮素及其配比对颠茄生长和氮代谢的影响
[韦悦1, 2 , 张翠平1, 2 , 郭双1, 2 , 卢克欢1, 2 , 吴能表1, 2  ]
]
]
|
|


第一作者:韦悦(1993-),女,重庆大足人,在读硕士生,主要从事药用植物次生代谢研究。E-mail:[email protected]
本研究采用砂培、浇灌营养液方法种植颠茄( Atropa belladonna),研究不同铵硝配比(0∶100、25∶75、50∶50、75∶25、100∶0)在不同处理时间(7、14、21、28 d)对颠茄干重、叶绿素和主要含氮化合物含量以及氮代谢关键酶活性的影响,为颠茄的合理施肥、科学种植提供理论依据。结果表明,颠茄叶片叶绿素、游离氨基酸(FAA)及可溶性蛋白(SP)含量随铵态氮比例增加而升高,其中叶绿素、FAA含量在铵硝比75∶25时最高,SP含量在铵硝比50∶50时最高,但这种影响在不同处理时间表现并不相同;整个处理期内,全硝营养下颠茄地上部和地下部干重明显高于铵硝混合处理及全铵处理,且叶片硝酸还原酶(NR)、谷氨酰胺合成酶(GS)活性较高,硝态氮含量也最高。综上,增加硝态氮含量能够促进颠茄干物质的积累以及NR和GS活性,合理补充铵态氮则能提高叶片叶绿素、FAA及SP含量,因此铵硝结合供氮且铵硝配比为25∶75更有利于颠茄的生长和氮素代谢。
To offer theoretical principles for rationally fertilizing and scientifically planting Atropa belladonna, the effects of N
颠茄(Atropa belladonna)为茄科(Solanaceae)颠茄属的多年生草本植物, 又名“ 野山茄” 、“ 颠茄草” , 是一种经济价值较大的药用植物, 其主要有效成分是莨菪碱(天仙子胺)、东莨菪碱(莨菪胺)和阿托品(颠茄碱)等托品烷类生物碱(tropane alkaloids, TAs)。莨菪碱和东莨菪碱已在临床上被广泛应用, 它们的作用主要是抗胆碱能神经传导或阻滞副交感神经[1]。已有研究以颠茄种子为外植体建立了颠茄快速繁殖体系, 为颠茄的培育提供了新方法[2]。随着毛状根培养技术的发展, 越来越多的学者开始以颠茄毛状根为材料研究其生物碱代谢的机制, 并克隆出了颠茄生物碱代谢途径中的关键酶基因[3, 4, 5], 极大地帮助人们了解颠茄的次生代谢特性。在多年种植经验和研究的基础之上, 杨吉福等[6]对颠茄的栽培技术作了总结, 为颠茄的田间管理提供指导。然而, 关于颠茄种植过程中矿质营养对颠茄生长和生理生化特性的影响却鲜有报道。
氮素是植物生长发育所必需的矿质元素, 也是植物体内蛋白质、核酸、叶绿素、酶、维生素、生物碱和一些植物激素等结构和功能大分子的重要组分。铵态氮(N
颠茄种子购买于湖南永州。将颗粒饱满的种子浸泡于50 mmol· L-1的赤霉素溶液中, 2 d后取出, 于暗培养箱内(温度25 ℃, 湿度60%), 平铺在湿润的滤纸上待萌发, 期间保持滤纸湿润。7 d后将萌发的种子播种到盛有充分混匀介质(珍珠岩∶ 蛭石=1∶ 2)的营养钵(12 cm× 13 cm)中, 每盆4株, 共300盆, 每7 d浇灌一次基本营养液, 培养30 d后进行铵态氮和硝态氮处理。基本营养液中大量元素采用Hoagland营养液配方, 微量元素采用Arnon营养液配方, pH 6.0[13]。所用试剂均为分析纯(AR)。
在总氮量(15 mmol· L-1)一致的条件下, 设置5个N
| 表1 不同形态氮素配比处理 Table 1 Treatments of proportion of different nitrogen forms |
1.3.1 干重的测定 颠茄生长30 d后开始处理, 并于处理后的7、14、21和28 d分别取样, 每处理随机取样6株。将植株洗净擦干, 其中3株置于烘箱中80 ℃烘干至恒重, 再分别称量其地上部和地下部干重; 其余植株用液氮速冻, -80 ℃保存备用。
1.3.2 叶绿素含量测定 叶绿素含量测定参照张宪政[15]的方法, 使用丙酮-乙醇混合液法。
1.3.3 氮代谢生理指标测定 可溶性蛋白(SP)含量采用考马斯亮蓝(G-250)染色法测定[16]; 游离氨基酸(FAA)含量采用茚三酮显色法测定[16]; 硝态氮含量采用水杨酸比色法测定[16]。硝酸还原酶(NR)活性按照南京建成生物工程研究所的试剂盒 (A096)方法进行测定。谷氨酰胺合成酶(GS)活性按照苏州科铭生物技术有限公司生产的谷氨酰胺合成酶测试盒进行测定, GS活力单位U定义为每克组织在每毫升反应体系中每分钟使540 nm下吸光值变化0.01。
所有指标均设置3个重复, 使用Microsoft Excel(2007)和SPSS 22.0对所有数据进行统计分析, 采用One-way ANOVA对数据进行单因素方差分析, 采用Duncan新复极差法比较相同处理不同处理时间下的差异和相同处理时间不同处理间的差异, 显著水平为0.05。利用Origin 8.0进行绘图。数据均以平均值± 标准误(X± SE)表示。
两种形态氮素及配比营养条件下颠茄的生长状况明显不同(图1)。氮素对颠茄茎叶和根部的生长有着相似的影响, 随着硝态氮比例的增加, 颠茄茎叶和根部干重都逐渐升高, 其中除7 d外, 其它处理时间地上部分茎叶干重在全硝营养(T1)下最高, 显著高于T3, T4和T5(P< 0.05); 在28 d时根部干重在铵硝比25∶ 75(T2)时最高, 显著高于其它处理。从整个处理期看来, T1、T2植株生长迅速, 它们的地上部干重在处理28 d时分别是处理7 d时的4.5和3.0倍, 地下部干重分别是处理7 d时的3.3和3.5倍; 而T4、T5在处理28 d时地上部干重仅分别是处理7 d时的1.9和1.8倍, 地下部干重增长也较少, 说明提高硝态氮的比例能促进颠茄的生长。
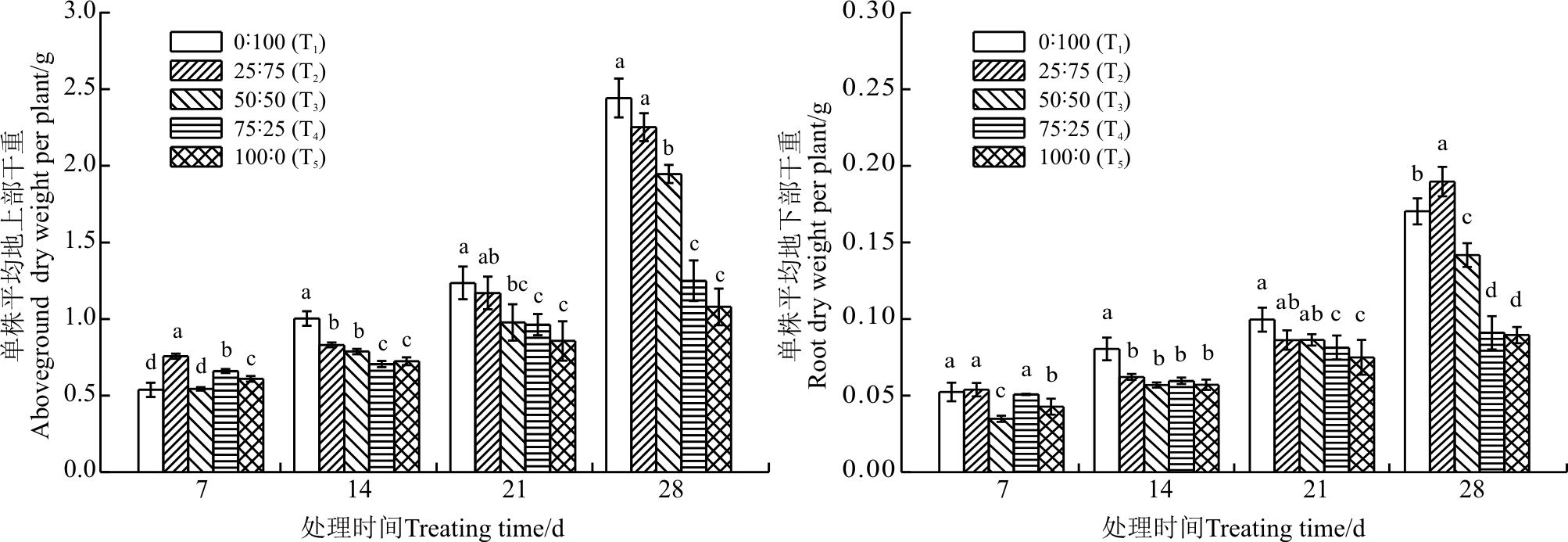 | 图1 铵态氮和硝态氮及其配比对颠茄地上部、地下部干重的影响 注:不同小写字母表示相同处理时间不同氮素处理间差异显著(P< 0.05)。T1、T2、T3、T4和T5分别代表营养液中N |
不同处理时期两种氮形态及配比下颠茄叶片中叶绿素含量变化不同(表2)。处理初期(7和14 d)T5叶绿素a、叶绿素b和总叶绿素含量最高, 其中叶绿素a和总叶绿素含量显著高于其它处理(P< 0.05), T1、T2和T4则紧随其后, T3的叶绿素a、叶绿素b及总叶绿素含量最低; 随着处理天数的增加, T2、T3、T4和T5叶绿素含量都呈上升趋势, 不同的是T1在处理21 d后开始下降, 而T2、T3和T4则持续上升, 处理28 d时显著升高。各处理间相比, T4在28 d时最高, 其中T4总叶绿素含量在处理28 d时超过T5, 达到3.685 mg· g-1, 说明处理初期全铵态氮供应比其它铵硝配比更有利于叶绿素的合成, 并且铵态氮的提升效果大于硝态氮, 而处理后期少量硝态氮(N
| 表2 氮素形态配比对颠茄叶片叶绿素含量的影响 Table 2 Effect of nitrogen form on chlorophyll content of Atropa belladonna leaves |
可溶性蛋白是植物体内主要的含氮化合物之一, 其含量对于研究植物氮代谢水平有重要参考价值。不同氮素形态处理下颠茄叶片可溶性蛋白含量变化明显(表3), 但是不同处理时间测定, 其变化规律不一致。处理7 d, T5叶片可溶性蛋白含量显著高于不同铵硝配比供氮组(T2、T3和T4)和全硝态氮T1(P< 0.05)。但随着处理天数的增加, 在21 d前T1可溶性蛋白含量呈下降趋势, T5可溶性蛋白含量没有显著变化, 直到处理28 d显著上升; 而T3、T4可溶性蛋白含量随着处理天数的增加则总体持续上升, 其中T3处理28 d时最高, 为1.048 mg· g-1 。说明全铵和全硝处理均能在短期(处理时间< 7 d)内能刺激可溶性蛋白的合成, 长期处于单一形态氮营养状况则不利于颠茄可溶性蛋白的积累; 相对于硝态氮, 铵态氮更能提高叶片可溶性蛋白的含量。
氮素在植物体内除了以结合态的蛋白质形式储存外, 还能以游离态的氨基酸形式存在, 游离氨基酸(FAA)在维持植物代谢平衡以及适应胁迫方面发挥着重要的作用。随处理时间增加各组FAA变化规律不同(表3), T1、T2的FAA含量先升高后降低; T3、T4、T5则表现出逐渐升高的趋势。随着处理液中铵态氮比例的增加叶片游离总氨基酸含量总体表现为升高, 其中T4处理的游离总氨基酸含量最高, 说明增加铵态氮含量有助于积累颠茄叶片中游离氨基酸。
硝态氮是能够被植物根部直接吸收的氮源[12], 其在植物体内的含量对于研究植物氮代谢机制有重要意义。处理7 d时全硝处理(T1)下颠茄叶片硝态氮含量是全铵处理(T5)下的2.1倍(表3), 处理28 d增加为6.1倍。此外, 相较处理7 d, 处理28 d后T1、T2、T5的硝态氮含量呈降低趋势。相反, T3处理硝态氮含量随处理时间增加总体呈现增加趋势, 但变化不显著(P> 0.05), T4则在处理28 d时硝态氮含量显著增加。说明在不同处理阶段不同铵硝配比对颠茄叶片硝态氮含量的影响都趋于一致, 叶片中硝态氮含量随着处理液中硝态氮比例的提高而增加, 而不同铵硝配比对颠茄硝态氮含量的影响随处理时间变化不同。
| 表3 氮素形态配比对颠茄叶片可溶性蛋白、游离氨基酸和硝态氮含量的影响 Table 3 Effect of nitrogen form on soluble proteins, free amino acids and nitrate contents of Atropa belladonna leaves |
两种氮素形态及配比对颠茄硝酸还原酶活性影响明显。整个处理时间内, 氮形态配比对硝配还原酶(NR)活性影响的变化规律都相似, 即NR活性随着处理液中硝态氮比例的增加而上升(表4)。处理28 d, 在铵硝比为25∶ 75(T2)条件下NR活性达到最高值27.927 μ mol· (mg· min)-1, 全铵营养下(T5)NR活性则下降为5.982 μ mol· (mg· min)-1, 显著低于T2处理(P< 0.05), 仅为最高值的21.4%。此外, 全硝营养下随着培养时间的增加NR活性先升高后降低, 说明即使提高硝态氮比例有助于增加颠茄叶片NR活性, 但从长期培养来看单一硝态氮供应并不利于颠茄植株对氮的同化。
相较于硝酸还原酶, 颠茄谷氨酰胺合成酶(GS)对两种氮素形态及配比变化的响应则不同, 随着硝态氮比例的减小GS活性呈现先减小后升高的趋势, 除处理21 d外, 其余处理时间下铵硝比50∶ 50(T3)为最低, 尤其是处理7 d时T3的叶片GS活性处理仅为3.175 μ mol· (mg· min)-1, 显著低于其它处理(P< 0.05), 而T1和T5在整个处理阶段内GS活性都保持较高水平。其中T1叶片GS活性在处理7 d时最高, 为15.556 μ mol· (mg· min)-1, 显著高于其余处理时间, 其余处理时间无显著变化(P> 0.05); T5处理组GS活性在处理28 d时最高, 为14.349 μ mol· (mg· min)-1。综上, 说明全铵和全硝态氮营养更能促进颠茄叶片GS活性。
| 表4 两种氮素形态及配比对颠茄硝酸还原酶、谷氨酰胺合成酶活性的影响 Table 4 Effect of nitrogen form on nitrate reductase and glutamine synthetase activities of Atropa belladonna leaves |
植物在长期进化过程中形成了对不同形态氮素的利用特性。有些植物更易于吸收铵态氮, 有些植物则偏好硝态氮, 另一些则在混合氮源中生长更好[17]。有研究报道单一硝态氮或高浓度铵态氮营养下草莓(Fragaria ananassa)的干重和鲜重都有所减少[18]。对桑树幼苗(Morus alba cv. Qinglong)的研究发现在单一形态氮营养下其植株高度、叶片数、叶片面积和根系长度均低于铵态氮和硝态氮配合施用[19]。芜菁(Brassica rapa)和芥菜(Brassica juncea)在单一铵态氮营养下的地上部干重和鲜重都较小, 叶面积和叶片数也减少[20]。根据植物在不同氮素形态营养下生长状况的好坏, 可将植物分为喜硝植物和喜铵植物[17]。本研究结果表明, 颠茄具备一定的喜硝特性, 在全硝营养下其地上部和地下部均能积累较多的干物质, 少量铵态氮(N
叶绿素是绿色植物叶绿体中的主要色素, 是植物光合作用中将光能转变为化学能并用于物质合成的关键物质, 其含量多少直接影响植物的生长。叶绿素a、叶绿素b都是含氮的化合物, 因此氮素的形态与叶绿素的合成密切相关。有研究指出, 铵硝等量供应时植物能够积累较多的叶绿素[25, 26]。然而本研究发现, 处理7和14 d时100%N
植物氮代谢包括初级氮同化、光呼吸产生的氮再同化和循环氮的再同化。其中以初级氮同化最为主要, 它由植物对氮素的吸收、硝态氮还原为氨以及氨同化为有机氮化物等过程组成, 硝酸还原酶(NR)和谷氨酰胺合成酶(GS)是这些过程中两个最重要的酶, 它们分别催化硝酸盐的还原和铵盐的同化[17]。氮素形态对NR和GS有很重要的调节作用[29]。对樱桃番茄[30](Lycopersivon esculentum)和掌叶半夏[31](Pinellia pedatisecta)的研究均指出NR活性与硝态氮含量正相关, 增加硝态氮的比例能够提高NR活性。对Vaccinium arboreum[32]的研究也发现硝态氮含量的增加与根系中NR活性的提高密切相关。本研究结论与上述研究结论一致, 随着处理液中硝态氮比例的增加颠茄叶片NR活性逐渐升高, 并分别在不同处理时间以铵硝比0∶ 100和25∶ 75较高, 且此时叶片硝态氮含量也较高。硝态氮不仅是NR作用的底物, 也是NR的诱导剂之一, 因此硝态氮对NR活性具有促进作用[33]。然而硝态氮对颠茄可溶性蛋白和FAA的积累却起到相反的作用, 这两种物质都随着硝态氮含量的减少而增加。GS则在单一硝态氮和单一铵态氮处理下均有较高的活性, 推测这种变化与氨的同化过程有关。全硝处理时由于NR活性的提高使大量的硝酸盐向铵盐转化, 相当于间接增加了GS作用的底物铵离子, 因此此时GS也具有较高的活性。在GS的作用下, 铵与谷氨酸结合生成了谷氨酰胺, 管氨酰胺又进一步转化为其它氨基酸, 这些氨基酸可以单独存在也可以用于合成蛋白质, 因此, 颠茄可溶性蛋白和游离氨基酸均在多铵处理下含量较高。但是, 也有研究认为, N
综上所述, 增加硝态氮含量能够促进颠茄干物质的积累以及NR和GS活性, 合理补充铵态氮则能提高叶片叶绿素、可溶性蛋白及游离氨基酸含量。因此铵硝结合供氮且铵硝配比为25∶ 75更有利于颠茄的生长和氮素代谢。此外, 这些生理特性也会进一步对颠茄次生代谢产生影响, 从而改变颠茄药用成分的含量。在实际生产中, 应综合考虑氮素对颠茄生理特性的影响, 或在不同时期调整氮素形态的供应以达到调节生长的目的。
Hu R F, Zhang G Q, Teng Z Y, Lin G Q.Effect of different nitrogens on activities of nitrate reductase, glutamine synthetase and seed protein contents in soybean cultivars.Journal of Northeast Agricultural University, 2012, 43(1):31-35.(in Chinese) (责任编辑 张瑾)
The authors have declared that no competing interests exist.
| [1] |
|
| [2] |
|
| [3] |
|
| [4] |
|
| [5] |
|
| [6] |
|
| [7] |
|
| [8] |
|
| [9] |
|
| [10] |
|
| [11] |
|
| [12] |
|
| [13] |
|
| [14] |
|
| [15] |
|
| [16] |
|
| [17] |
|
| [18] |
|
| [19] |
|
| [20] |
|
| [21] |
|
| [22] |
|
| [23] |
|
| [24] |
|
| [25] |
|
| [26] |
|
| [27] |
|
| [28] |
|
| [29] |
|
| [30] |
|
| [31] |
|
| [32] |
|
| [33] |
|
| [34] |
|
两种形态氮素及其配比对颠茄生长和氮代谢的影响
韦悦
Effect of nitrogen form on growth and nitrogen metabolism of Atropa belladonna
Wei Yue