{kind=link}
{kind=link}
两种藓类植物对光强的适应性差异
[姬明飞1  , 韩鸿基
, 韩鸿基2 ]
, 韩鸿基|
|


以宝天曼自然保护区两种藓类物种大羽藓( Thuidium cymbifolium)和无边提灯藓( Mnium immarginatum)为研究对象,测定和分析其光合生理特征对于不同光强的响应,探讨藓类植物光适应能力差异的原因,以期为更加合理地保护、利用苔藓植物资源提供理论依据。结果表明,无边提灯藓的表观量子效率(AQY)、光饱和点(LSP )以及光补偿点(LCP)都显著低于大羽藓( P<0.05),但是大羽藓最大光合速率(Amax)显著高于无边提灯藓,说明无边提灯藓对于弱光生境的适应能力相对较强。进一步从光合参数-光响应过程中分析发现,两种藓类植物的胞间CO2浓度(Ci)和蒸腾速率(Tr)随光照强度变化的趋势基本一致,但大羽藓的水分利用效率(WUE)和气孔导度(Gs)高于无边提灯藓,这从另一个侧面反映出大羽藓独特的光适应策略。综上所述,气孔导度水平的差异可能是最终导致两种藓类植物对于不同光环境适应性不同的原因。
To explore the adaptive mechanisms of bryophyte plants in Baotianman Nature Reserve, our research determined the photosynthetic characteristics of Thuidium cymbifolium and Mnium immarginatum and revealed a trend of variation with light gradients. Our results showed that M. immarginatum had lower apparent quantum efficiency (AQY), light saturation point (LSP), and light compensation point (LCP) values than T. cymbifolium, indicating that M. immarginatum has a greater ability to adapt to low-light conditions; however, the maximum net photosynthetic rate (Amax) value for T. cymbifolium suggested that it may employ a unique light adaptation strategy. When considering the photosynthetic characteristics of these two species, T. cymbifolium exhibited higher values of water use efficiency (WUE) and stomatal conductance (Gs), which were quite different from intercellular CO2 concentration (Ci) and transpiration rate (Tr). Overall, the results confirmed the hypothesis that a difference in Gs between these two species resulted in adaptation to the environment and represents a survival strategy.
光合作用作为植物最重要的生理过程, 解析净光合速率(Pn)、气孔导度(Gs)、蒸腾速率(Tr)、水分利用效率(WUE)等生理生态指标对环境变化的响应特征, 研究光合作用有助于了解植物的生长发育规律, 探寻植物对不同环境的适应机理[1, 2]。已有的研究表明, 通过研究光合-光响应曲线模型, 能够更加直观地了解植物对不同光环境的适应过程[3, 4]。关于光响应曲线数据的拟合方法有许多种[5, 6, 7, 8], 其中由于非直角双曲线拟合参数K(shape parameter)与叶片厚度和单位面积叶绿素含量存在密切关系而更具生物学意义[9, 10], 因而非直角双曲线拟合方法被广泛用于生理学和植物生理生态学的研究中[11]。
苔藓植物作为一个独特的植物类群, 在自然界中扮演着重要的角色。因其特殊的结构和生理生态学特性, 苔藓植物往往对干旱、高温、低温、强光辐射等逆境胁迫具有较强的适应性[12, 13, 14]。长期以来, 人们在苔藓植物的元素含量[15]、细胞超微结构、光合色素、渗透调节物(脯氨酸、可溶性糖、游离氨基酸等)等苔藓植物逆境适应机制方面进行了大量研究工作[16]。然而, 关于苔藓植物对于不同光强的适应性研究鲜有报道[17]。因此, 本研究以宝天曼自然保护区优势苔藓植物大羽藓(Thuidium cymbifolium)和无边提灯藓(Mnium immarginatum)为研究对象, 采用非直角双曲线拟合方程, 对宝天曼自然保护区广布的两种藓类植物的光合-光响应曲线进行拟合分析, 比较两种藓类植物的光合-光响应特征, 探讨其对不同光环境的适应机制, 以期为更加合理地保护、利用苔藓植物提供理论依据。
宝天曼自然保护区位于河南省西南部。地理坐标为33° 25'~33° 33' N, 111° 53'~112° 00' E。地处秦岭东段, 伏牛山南坡, 属北亚热带向南暖温带过渡区域。森林植被覆盖率95.4%, 群落垂直分布明显, 区内海拔相对高度差1 330 m。年均气温15.1 ℃, 1月平均气温1.5 ℃, 7月平均气温27.8 ℃; 年均降水量885.6 mm, 年均蒸发量991.6 mm, 年均相对湿度为68%。
本研究于2013年8月开展, 采用便携式光合测定系统(LI-6400, Li-Cor)进行光合速率测定。测量时, 选取当年生、成熟的绿色组织, 每个种选取3株个体, 鉴于苔藓植物较为低矮这种特殊的形态, 测定时流速设定为300 μ mol· mol-1。在测定的过程中, 尽量选取相同的时间段进行测定, 保持外界的环境一致性, 一般情况下CO2浓度控制在400 μ mol· L-1, 叶片的温度基本控制在28 ℃, 相对湿度为50%。光照强度设定分为0、40、80、150、200、400、600、800和1 200 μ mol· (m2· s)-1, 测定按照光照强度从高到低的顺序进行。
采用非直角双曲线模型拟合测得的光响应曲线[6], 其原理是根据下面所示的公式。
A(I)=
式中:A, 光合速率; I, 光量子通量密度(PPFD); Ø , 表观量子效率; Amax, 光饱和点时同化速率; Rd, 暗呼吸速率; θ , 曲线的曲率K[6])。通过这个模型, 可以计算出光饱和点(LSP)、光补偿点(LCP)、表观量子效率(AQY)和水分利用效率(WUE)。
所有的数据分析在SPSS和R语言境下进行分析。其中, 光曲线拟合利用SPSS(SPSS, Inc., Chicago, IL, USA), 具体参照参考文献[25]的拟合方法; 光响应的差异研究采取R语言环境(R Development Core Team, 2008)的lme分析包功能进行混合线性模型(Mixed Model Regression)分析。
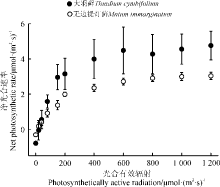
随着光强增加, 两种藓类植物都表现出逐渐上升到稳定的趋势。大羽藓具有较高的Amax和LSP, 分别为5.730和376.866 μ mol· (m2· s)-1(表1)。此外, LCP是植物利用弱光能力大小的指标, 两种藓类植物的光补偿点相对较低, 且大羽藓的LCP[20.862 μ mol· (m2· s)-1)]略高于无边提灯藓[18.027 μ mol· (m2· s)-1)], 说明两种藓类植物可能对弱光条件具有一定的适应能力, 且无边提灯藓可能较强(表1、图1)。
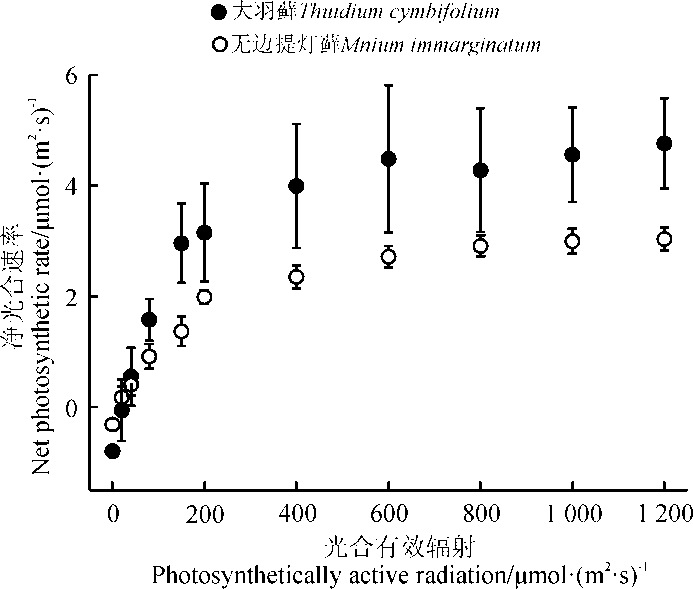 | 图1 两种藓类植物光合响应曲线Fig. 1 Photosynthesis-light response curves of two moss species |
两种藓类植物的AQY具有一定的差别, 大羽藓高于无边提灯藓(表1)。两种藓类植物整体上在弱光环境具有较好的适应性, 且大羽藓具有较高的光能转化效率。方差分析结果显示(表2), 光合-光响应曲线拟合参数除LCP之外, 其余指标种间差异显著(P< 0.05)。
| 表1 两种藓类植物非直角双曲线拟合的光合参数 Table 1 Photosynthetic parameters of two mass species from non-rectangle hyperbola model |
| 表2 光曲线拟合值在种间(S)以及个体间(I)的方差分析 Table 2 Variance components of the estimated parameters of the photosynthetic light-response curves between species and among individuals |
本研究中两种藓类植物中的Rd具有明显的差异, 大羽藓[0.401 μ mol· (m2· s)-1]高于无边提灯藓[0.191 μ mol· (m2· s)-1]。Rd作为一个与植物叶片生理活性和温度密切相关的指标, 植物生长的环境条件和其自身生理适应机制都会对光响应特征参数产生很大的影响。因此, 究其原因, 还需进一步研究两种藓类植物叶片的生理活性。
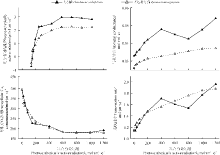
随着光强的增加, 两种藓类植物的Gs和Tr表现出不断上升的趋势, 但是大羽藓在光强为400~800 μ mol· (m2· s)-1表现出一定的下降趋势(图2)。两种藓类植物的Ci随着PAR的增加和Amax的增大而减小, 到PAR大约达到600 μ mol· (m2· s)-1时, 两种藓类植物的Ci降低至150~200 mol· (m2· s)-1, 基本维持平衡。除此之外, 两种藓类植物在PAR增大的初始阶段都有一个大量消耗CO2的过程。产生这样的现象, 可能是在光照增强的初期, Gs值还比较小, 导致藓类植物吸收外界CO2量不能满足光合过程的需求, 最终使得Ci明显下降。
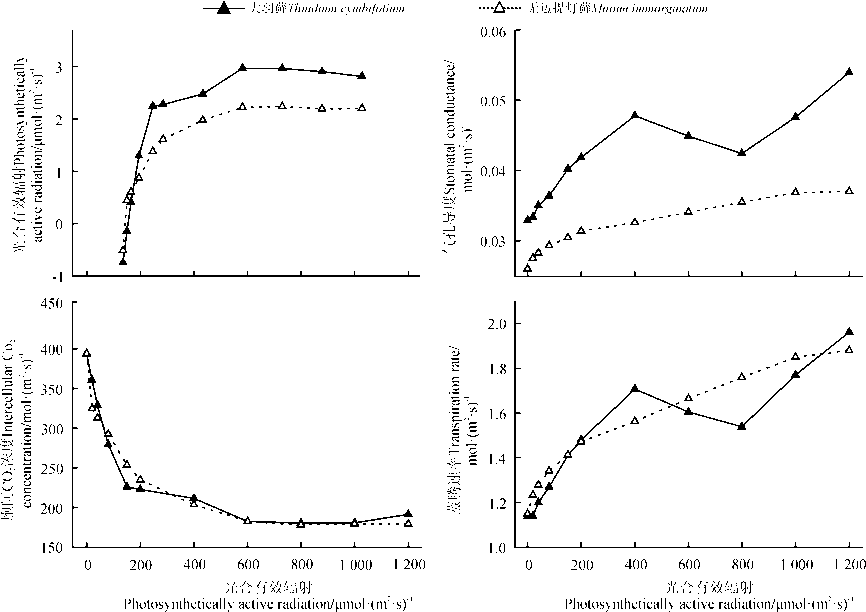 | 图2 4种光合参数的光响应过程Fig. 2 The responses of four photosynthetic parameters to light conditions |
两种藓类植物的Tr始终随着PAR的增加而升高, 并在PAR为400~600 μ mol· (m2· s)-1时, 有明显的下降趋势。同时, 两种材料的WUE表现出近似的变化趋势。WUE在起始阶段随着PAR的增加而逐渐上升, 当PAR达到600 μ mol· (m2· s)-1时, 两种藓类植物的WUE达到最大水分利用效率。此后, 随着PAR的升高, 两种藓类植物的WUE基本不再变化, 总体上一直维持在一个较高的水平。
通过对宝天曼自然保护区内两种主要藓类植物在不同光强下的适应性差异进行研究, 结果表明, 两种藓类植物在光能利用方面存在显著差异。光合-光响应曲线能准确地反映出植物对于光强的适应性[17, 18]。光补偿点是植物对于弱光利用能力大小的一个重要指标, 该指标表示植物在弱光条件下进行有机物积累的多少, 是植物耐阴性的主要参数[19]。结合前人对于附生地衣(Epiphytic lichens)的光合特征研究结果, 本研究的光补偿点略高于阴生类地衣[20], 这说明这两种藓类植物能较好地适应弱光环境, 不易发生光抑制现象。AQY作为弱光条件下植物对光的吸收、转换和利用能力强弱的重要指标[21]。由于植物和环境之间较为复杂的关系, 大部分研究认为AQY的经验值一般都在0.04~0.07为正常[22, 23]。本研究中AQY相对较低, 从另外一方面也证明, 林下弱光条件可能促使两种藓类植物对于光能的适应机制产生分化, 并形成了稳定且可以遗传的生存策略, 从而导致两种苔藓植物的光响应曲线拟合出的指标在种间存在较大的差异。这些结果也初步肯定了植物功能性状是植物自身对于环境长期适应而形成自身稳定遗传的生存策略[24, 25, 26]。
一般认为, 气孔是植物叶片与大气进行气体交换的主要通道, 并随着外界光强变化, 气孔开闭程度不同, 从而直接影响光合作用和蒸腾作用, 同时也对WUE产生间接影响[11]。已有的研究表明, 如果光合有效辐射超过饱和点, Gs上升而Ci浓度下降, 植物的净光合速率下降的原因可能是强光下的光抑制而非气孔因素导致[27]。本研究结果略有不同, Gs的变化没有直接导致两个物种的Ci浓度发生明显差异, 但是WUE产生明显变化。结合Gs的变化趋势, 推测可能是因为非光因素导致气孔的暂时性开放, 造成大羽藓的Ci暂时降低, 对于无边提灯藓来说, 其较低的气孔导度水平, 有可能是导致两个物种光能适应差异的主要导原因[28, 29]。
综上所述, 利用非直角双曲线可以在一定程度上反映藓类植物对于不同光环境的适应机制, 但是影响藓类植物气孔开闭的机制以及与环境因子的关系还有待于进一步的研究。此外, 以上的研究结果只是基于优势种得出的结论, 还需要更多其它藓类植物的研究进行补充。
The authors have declared that no competing interests exist.
| [1] |
|
| [2] |
|
| [3] |
|
| [4] |
|
| [5] |
|
| [6] |
|
| [7] |
|
| [8] |
|
| [9] |
|
| [10] |
|
| [11] |
|
| [12] |
|
| [13] |
|
| [14] |
|
| [15] |
|
| [16] |
|
| [17] |
|
| [18] |
|
| [19] |
|
| [20] |
|
| [21] |
|
| [22] |
|
| [23] |
|
| [24] |
|
| [25] |
|
| [26] |
|
| [27] |
|
| [28] |
|
| [29] |
|
两种藓类植物对光强的适应性差异
姬明飞, 韩鸿基
The study of photosynthetic characteristics of two moss species under different light conditions
Ji Ming-fei, Han Hong-ji