
{kind=link}
白花草木樨EST-SSR标记的开发与筛选
[剡转转, 任艳, 吴凡, 骆凯, 张代玉, 闫启, 张宇飞, 赵玉凤, 张吉宇 ]
]
]
|
|


第一作者:剡转转(1994-),女,甘肃静宁人,在读硕士生,主要从事牧草种质资源创新与利用。E-mail:[email protected]
草木樨( Melilotus)是重要的豆科牧草之一,然而草木樨分子标记匮乏,限制了草木樨种质资源的开发与利用。本研究以白花草木樨( M. albus)转录组数据为基础,设计了18 182对草木樨EST-SSR引物,并对所开发的EST-SSR引物进行筛选。通过PCR扩增从550对EST-SSR引物中筛选得到206对白花草木樨多态性引物,共检测出679个等位基因,平均每对引物检测出2.888个基因位点。多态信息含量PIC的分布范围为0.239~0.855,平均值为0.468。206对多态性引物Nei’s基因多样性指数(H)和Shannon信息指数(I)的平均值分别为0.169和0.239。本研究开发的EST-SSR丰富了草木樨属的分子标记,为研究种质资源的遗传多样性和分子辅助育种奠定了基础。
Melilotus is one of the most important legume forages, however, the lack of molecular markers has limited the development and utilization of Melilotus germplasm resources in breeding programs. In this study, 18 182 EST-SSR primers were developed and screened based on transcriptome data from M. albus. A total of 206 primer pairs from M. albus were found to be polymorphic from a total of 550 pairs of EST-SSR primers screened. A total of 679 alleles were detected with an average of 2.888 alleles per locus. The polymorphism information content (PIC) values ranged from 0.239 to 0.855, with an average of 0.468. Nei’s gene diversity and Shannon’s information index for 206 pairs of polymorphic primers were 0.169, and 0.239, respectively. The EST-SSRs developed in this study enriches the EST-SSR molecular markers of Melilotus, which will serve as the foundation for exploring the genetic diversity of germplasm resources for marker-assisted breeding.
草木樨(Melilotus spp.)为豆科一年生或二年生自花授粉或异花授粉二倍体植物[1, 2], 全世界约有19种, 主要分布于欧洲地中海区域、东欧和亚洲, 在我国主要分布于东北、华南、西南。白花草木樨(M. albus)为该属生产利用常见种之一, 自交亲和率约为49%[2], 具有适应性广、抗逆性强[3]、繁殖速度快、种子产量高以及固氮能力强等优良特性[4], 是仅次于苜蓿(Medicago sativa)的优良豆科牧草。作为豆科牧草, 草木樨的固氮能力高, 有利于草田轮作, 因此, 发展草木樨是提高农作物产量的一项有效措施[5]。近年来, 我国越来越重视草牧业的发展, 提出将粮食作物、经济作物的二元结构调整为粮食作物、经济作物、饲料作物的三元结构, 通过种植优质高产饲草, 推进粮食作物种植向饲草料作物种植方向转变[6], 草木樨是粮改饲的主要草种之一。
简单序列重复(simple sequence repeats, SSR)标记是一类由几个核苷酸(一般为1~6个)为重复单位组成的长达几十个核苷酸的串联重复序列。因具有多态性高、位点特异性、突变率高、共显性、操作简单等优点[7, 8, 9, 10], 近年来已经快速发展成为国内外广泛应用的方法[11], 利用分子标记技术来研究作物种质资源间的遗传变异也越来越受到人们重视, SSR标记还可以用来鉴别品种间的亲缘关系[12]。EST-SSR(express sequence taget-simple sequence repeat)是基于表达序列标签开发的微卫星分子标记, 又称表达序列标签-简单序列重复。由于EST-SSR来源于基因组中的转录区域, 且在同源基因中含有保守序列, 因此, 与基因组SSR相比, EST-SSR具有在植物物种之间可转移性的优点。目前, EST-SSR分子标记已在多个物种中得到了应用。Zhou等[13]利用EST-SSR对老芒麦(Elymus sibiricus)的遗传多样性进行了研究, 推动了披碱草属植物分子遗传多样性的研究。EST-SSR分子标记已经在油菜(Brassica campestris)[14]、茶(Camellia sinensis)树[15]等作物和紫花苜蓿[16]、高丹草(Sorghum bicolor× S. sudanenes)[17]等牧草中得到了应用。
草木樨属的研究大多集中在栽培技术[18]、化学成分[19]等方面。骆凯等[20]对从国内外引进的19份二年生草木樨种质进行了农艺性状和品质的初步评价; Di等[21]对整个草木樨属的形态学以及分子系统进化进行了研究, 证实草木樨属是单系统遗传, 与苜蓿、三叶草(Trifolium repens)的亲缘关系最近。然而目前关于草木樨分子标记开发的研究报道较少, 可用于研究草木樨属植物的遗传工具和资源非常有限。2007年Winton等[22]开发了9对草木樨基因组SSR标记研究了草木樨在阿拉斯加洲入侵的起源。随后Wu等[11]采用18对SSR引物研究了草木樨属18个种的遗传多样性。本研究利用生物信息学工具, 从白花草木樨转录组数据中开发设计大规模的EST-SSR分子标记, 进一步筛选多态性高标记, 以期为草木樨属植物的遗传资源开发与利用奠定基础。
白花草木樨的5份种质来自我国国家牧草种质资源库(北京)(表1)。其中, ZXY06P-1732和 ZXY05P-983来自俄罗斯, HB2009-153来自中国信阳, 中畜-1226和ZXY07P-3150来源地不详。
| 表1 供试材料及来源信息 Table 1 Origin information of the tested Melilotus germplasm |
1.2.1 EST-SSR引物的开发与设计 由诺和公司完成白花草木樨转录组测序数据已提交到NCBI(http://www.ncbi.nlm.nih.gov/Traces/sra), 采用MISA(1.0版)对Unigene进行SSR检测, 识别标准为:包含1、2、3、4、5、6个碱基重复单元的EST-SSR分子标记的最小重复数分别为10、6、5、5、5、5。通过两个引物之间不发生互补, 特别是在引物3'端, 即使无法避免, 其3'端互补碱基也应不大于两个碱基的筛选原则, 利用软件Primer3[23, 24](默认参数)设计EST-SSR引物。
1.2.2 DNA的提取 每种材料取18个单株的新鲜叶片混合, 用SDS法[25]提取叶片的总DNA, 将提取出来的DNA用琼脂糖凝胶电泳检测其质量, 对合格样品加ddH2O稀释到50 ng· μ L-1, 置于-20 ℃冰箱中保存。
1.2.3 引物筛选、PCR扩增及电泳 转录组测序获得的18 182对EST-SSR引物, 根据引物的碱基重复类型(除一碱基之外)、片段长度(150~200 bp)、退火温度(55~60 ℃)等条件筛选得到550对EST-SSR引物用于本试验。PCR反应体系总体积为10 μ L, 含1.0 μ L模板DNA, 4 μ mol· L-1正反引物各1 μ L, 0.1 μ LDNA聚合酶, 4.9 μ L 2 × Reaction Mix, 2 μ L ddH2O。初步筛选反应程序:94 ℃预变性3 min; 94 ℃变性30 s, 引物最适退火温度退火30 s, 72 ℃延伸30 s, 35个循环; 72 ℃延伸7 min; 4 ℃保存。扩增产物用6.0%的变性聚丙烯酰胺凝胶电泳分离, 银染。本试验使用DL500DNA Marker。
通过人工读带, 根据扩增条带在相对迁移位置的有无, 记数为“ 1” 或“ 0” , 建立原始数据矩阵, 统计每个EST-SSR位点的等位基因数, 计算引物的多态信息含量PIC[26]以及期望杂合度(He)。
PIC=1-∑
式中:Pi指第i个等位基因出现的频率。
He=1-
用Popgen32软件统计分析Nei’ s多样性指数和Shannon信息指数。
通过SSR识别工具软件MISA(1.0版)对得到的功能基因进行位点识别后, 从15 260个功能基因中共识别出了19 263个潜在的EST-SSR位点。EST-SSR引物的设计由Primer3软件完成, 设计得到18 182对EST-SSR引物。其中单碱基的有11 398对, 二碱基的有2 986对, 三碱基的有3 461对, 四碱基的有299对, 五碱基的有29对, 六碱基的有9对。进一步根据碱基重复类型、预期片段大小和退火温度等条件来挑选, 共有符合要求的550对引物EST-SSR用于进一步筛选, 其中二碱基的有136对, 三碱基的有317对, 四碱基的有84对, 五碱基的有12对, 六碱基的有1对(表2)。
| 表2 EST-SSR引物的来源及碱基重复类型 Table 2 The origin and variation type of the EST-SSR primers |
利用白花草木樨材料对550对EST-SSR引物进行筛选验证, 结果表明, 共有351对引物能够扩增出条带, 而剩余的199对引物在多个退火温度下都没有检测出PCR扩增产物。在能够成功扩增出条带的草木樨EST-SSR引物中, 共有290对引物获得条带清晰、大小合适的扩增产物, 而剩余的61对引物扩增出的PCR产物条带大小都大于或小于引物的预期片段大小。经过计算各个引物的多态性指标, 在能够扩增出引物预期片段大小的草木樨EST-SSR引物中, 206对引物表现出多态性(图1), 而剩余的84对引物未表现出多态性(表3)。206对草木樨EST-SSR引物共检测出679个等位基因, 变幅为2~6个, 平均每对引物检测出2.888个基因位点, 其中c9516_g1和c9897_g检测到的等位变异数最多, 为6个。期望杂合度的变化范围为0.278~0.861, 平均值为0.536。多态信息含量PIC的分布范围为0.239~0.855, 平均值为0.468(表3)。
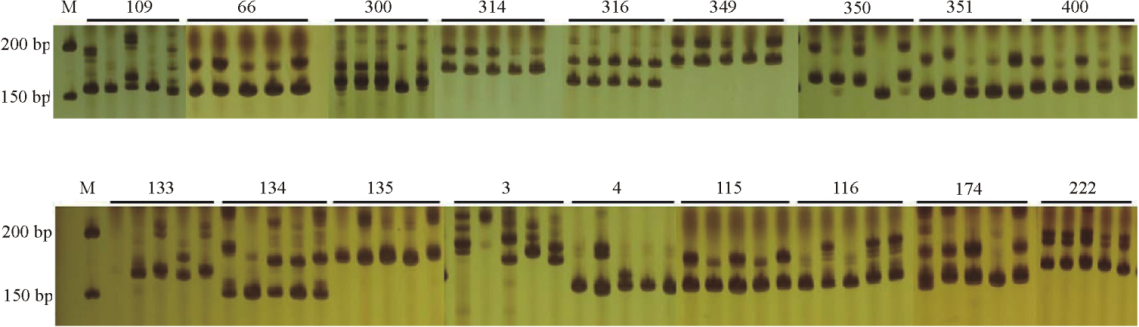 | 图1 部分引物对5份白花草木樨种质的扩增结果 注:图中引物编号与表3相同。M, Marker, DL500DNA。Fig. 1 Amplification products from five Melilotus albus accessions using select primer pairs Note: Numbers of the primers refer to Table 3. M, Marker, DL500DNA. |
用550对EST-SSR引物对5份白花草木樨种质进行扩增, 结果表明, 多态性EST-SSR标记位点的观测等位基因数(Na)、期望等位基因数(Ne)、Nei’ s基因多样性指数(H)和Shannon信息指数(I)的平均值分别为1.810、1.541、0.316和0.466。
白花草木樨5份种质的遗传距离介于0.455~0.580, Pop1、Pop3和Pop1与Pop4的遗传距离最大, 均为0.580; Pop1与Pop2遗传距离最小, 为0.455; 5份种质的遗传一致度介于0.550~0.634, Pop1与Pop2之间的遗传一致度最大, 为0.634(表4), 表明这两份种质间的遗传相似性最高。Pop1、Pop3和Pop1与Pop4的遗传一致度最小, 均为0.556, 表明这两份种质间的遗传相似性最低。
| 表4 5份种质的Nei’ s遗传一致度和遗传距离 Table 4 Nei’ s genetic similarity coefficient and genetic distance between five accessions |
分子标记是作物分子遗传育种研究的重要工具之一。在众多的分子标记中, SSR已被广泛应用于种质遗传多样性的鉴定、分析以及种质遗传图谱的构建等研究中[27]。SSR分子标记已经在藜麦(Chenopodium quinoa)[28]、小麦(Triticum aestivum)[29]等主要农作物的种质资源研究中得到了广泛应用, 但在草木樨等豆科作物上, 由于开发的分子标记较少, 检测到的SSR分子水平的多态性较少, 其应用受到了限制。近年来, 通过生物信息学手段利用公共数据库中丰富的基因组和转录组序列发展分子标记成为开发SSR及其它类型分子标记的重要途径[30, 31, 32]。本研究通过NCBI数据库白花草木樨转录组测序设计出18 182对EST-SSR引物。并通过草木樨5份种质对550对EST-SSR引物进行筛选, 共检测出679个等位基因, 变幅为2~6个, 平均每个位点2.888个。多态性信息含量PIC的分布范围为0.239~0.855, 平均值为0.468。Nei’ s基因多样性指数、Shannon信息指数的平均值分别为0.169、0.239。陈立强和师尚礼[16]用15对SSR引物在42份紫花苜蓿材料中共检测到231条扩增带, 其中163条具有多态性, Nei’ s基因多样性指数和Shannon信息指数(I)的平均值分别为0.210 0和0.326 2。Wu等[11]利用18对SSR引物在草木樨18个种共50份种质中共检测到287个等位基因, 平均每个位点15.94个, 多态信息含量PIC 的分布范围为0.71~0.93, 平均值为0.87。Winton等[22]开发出的9对草木樨基因组SSR在白花草木樨29个种质中共检测到36个等位基因, 平均每个位点4个。本研究所表现出的引物多态信指数相对较低, 原因可能有:一是EST编码功能基因, 序列保守性高, EST-SSR的多态性低于基因组SSR[33, 34]。二是本研究所选的材料是草木樨属种内的材料, 所选材料间的亲缘关系较近, 不易得到多态性高的引物。
本研究从转录组测序获得18 182对EST-SSR引物, 根据引物的碱基重复类型、片段长度、退火温度等条件筛选得到550对EST-SSR引物, 再次筛选, 共得到290对有扩增产物的引物, 其中206对引物具有多态性。结果表明, 白花草木樨EST-SSR含有丰富的SSR位点, 本研究开发的EST-SSR丰富了草木樨属EST-SSR分子标记, 为研究种质资源的遗传多样性和分子辅助育种奠定了基础。
The authors have declared that no competing interests exist.
| [1] |
|
| [2] |
|
| [3] |
|
| [4] |
|
| [5] |
|
| [6] |
|
| [7] |
|
| [8] |
|
| [9] |
|
| [10] |
|
| [11] |
|
| [12] |
|
| [13] |
|
| [14] |
|
| [15] |
|
| [16] |
|
| [17] |
|
| [18] |
|
| [19] |
|
| [20] |
|
| [21] |
|
| [22] |
|
| [23] |
|
| [24] |
|
| [25] |
|
| [26] |
|
| [27] |
|
| [28] |
|
| [29] |
|
| [30] |
|
| [31] |
|
| [32] |
|
| [33] |
|
| [34] |
|
白花草木樨EST-SSR标记的开发与筛选
剡转转, 任艳, 吴凡, 骆凯, 张代玉, 闫启, 张宇飞, 赵玉凤, 张吉宇
The development and screening of EST-SSR markers in Melilotus albus
Yan Zhuan-zhuan, Ren Yan, Wu Fan, Luo Kai, Zhang Dai-yu, Yan Qi, Zhang Yu-fei, Zhao Yu-feng, Zhang Ji-yu