{kind=link}
{kind=link}
{kind=link}
稻-麦连作和稻-休耕农田植物物种多样性的比较
引用本文
吴芸紫, 刘章勇, 蒋哲, 杨梅. 稻-麦连作和稻-休耕农田植物物种多样性的比较. 草业科学, 2017,34(5):1090-1099
Wu Yun-zi, Liu Zhang-yong, Jiang Zhe, Yang Mei. Comparison of plant species diversity in continuous rice-wheat and rice-fallow systems. Pratacultural Science,2017,34(5): 1090-1099
Permissions
Wu Yun-zi, Liu Zhang-yong, Jiang Zhe, Yang Mei. Comparison of plant species diversity in continuous rice-wheat and rice-fallow systems. Pratacultural Science,2017,34(5): 1090-1099
Copyright©2017, 《草业科学》编辑部
《草业科学》编辑部
稻-麦连作和稻-休耕农田植物物种多样性的比较
第一作者:吴芸紫(1991-),女,湖北咸宁人,在读硕士生,主要从事农业生物多样性研究。E-mail:[email protected]
摘要
为揭示休耕对农田生态系统植物群落和物种多样性的影响,以稻-麦连作和稻-休耕系统为研究对象,研究田内、田埂和沟渠3种生境的物种组成和多样性特征。结果表明,1)稻-麦连作系统植物由28科58属64种组成,多年生植物占39.1%;休耕系统植物由32科68属76种组成,多年生植物占52.6%;2)休耕显著提高了物种丰富度( P=0.021)和Shannon-Weiner多样性指数( P=0.006),但是稻-麦连作和稻-休耕系统之间的Simpson优势度指数和Pielou均匀度指数没有显著差异( P=0 .318, P=0.844);3)无论稻-麦连作还是稻-休耕系统,田埂生境中各多样性指数最高;但在休耕系统中,田埂和田内各多样性指数均没有显著差异;4)样方去趋势对应分析(detrended correspondence analysis, DCA)排序明确揭示了连作和休耕系统样地基本分离,且休耕期田内和田埂植物群落组成相似。休耕可降低作物生境和非作物生境的植物组成差异,促进农田生态系统植物物种多样性。
关键词:
稻-麦连作; 稻-休耕; DCA排序; 物种丰富度; 优势度指数; 物种多样性指数; 生境
中图分类号:S344.1
文献标志码:A
文章编号:1001-0629(2017)05-1090-10
doi: 10.11829/j.issn.1001-0629.2016-0554
Comparison of plant species diversity in continuous rice-wheat and rice-fallow systems
Abstract
To explore the effect of fallow on weed community and species diversity in farm ecosystem, species composition and diversity were investigated in the interior, field bund, and drainage ditch of continuous rice-wheat and rice-fallow systems. The results showed that, 1) 64 species (58 genera belonging to 28 families) were distributed in continuous rice-wheat system with 39.1% perennial plants, while 76 species (32 families belonging to 68 genera) were distributed in rice-fallow system with 52.6% perennial plants; 2) Fallow significantly increased species richness ( P=0.021)and Shannon-Wiener diversity index( P=0.006), but the difference of Simpson dominance index and Pielou evenness ( P=0.318)index were not significant( P=0.844), between the two systems; 3) The species index in field bund were higher than that in interior and drainage ditch of both systems, which showed the importance of non-crop habitats for maintaining biodiversity, but no significant differences were found between the margin and interior of rice-fallow system; 4) The results of DCA clearly reflected the distribution range of various community types. Almost all rice-wheat communities were separated from rice-fallow communities, and community composition were similar between interior and field bund in the fallow period. These results reveal that fallow can reduce the difference of species composition between crop and non-crop habitats and effectively improve the plant biodiversity level of farm ecosystem.
Keyword:
continuous rice-wheat system; rice-fallow system; DCA ordination; species richness; dominance index; diversity index; habitats
农田植物多样性对于增加天敌数量, 防止土壤侵蚀, 促进养分积累与循环, 调节农田小气候等农田生态系统服务功能的维持具有重要作用[1, 2]。近年来, 关于农田植物多样性特征及其影响因素的研究日益受到重视[3, 4], 农田管理方式对农田植物多样性有重要影响, 包括农业化学品的使用[5, 6, 7, 8, 9]、种植制度(如, 作物配置、熟制和种植方式)[10, 11]以及耕作方式(如, 免耕、旋耕、深松)等[12, 13, 14, 15, 16]。农田管理方式直接关系到农田生态环境、土壤养分和土壤水分等的改变, 进一步影响农田植物之间的竞争关系, 导致植物群落组成和多样性的不同[3, 15]。尽管如此, 国内外研究多集中在化肥、除草剂、轮作和保护性耕作等对农田植物群落的影响[5, 6, 7, 8, 9, 10, 11, 12, 13, 14, 15, 16], 针对作物-休耕制度对植物多样性影响的研究仍少见报道。
休耕是提高耕地质量且成本低、见效快的有效途径, 它是指在可耕、可种的季节不耕田, 不种植作物的一种耕作方式, 可分为季休、年轮休、长休等[17]。休耕有利于缓解作物期带来的负面效应, 提高土壤肥力和降低土壤侵蚀风险, 提高物种多样性[18, 19, 20, 21]。我国传统农业很重视休耕, 国外则常通过休耕恢复牧场和保护农田鸟类[22]。稻-麦两熟和稻-休耕一熟是目前我国长江中下游稻作区主要的种植制度。本研究通过比较连续3年稻-麦连作和稻-休耕系统的植物物种多样性和群落结构, 以期阐明休耕对植物多样性和植物群落组成的影响, 揭示其规律为农田管理及农田植物多样性保护提供科学依据。
1 研究区概况与研究方法
1.1 研究区概况
试验地位于荆州市太湖农场(30° 24' N, 111° 54' E)。该区域是江汉平原粮食主产区腹地, 地处亚热带季风气候区, 年日照时数1 800~2 000 h, 年平均气温15.9~16.6 ℃, 年无霜期242~263 d, 多数年降水量在1 100~1 300 mm, 4月-10月降水量占全年80%。调查区各样地作物期的农事操作基本相同, 各样地面积在860~1 240 m2, 平均面积为1 050 m2。
1.2 研究方法
1.2.1 调查方法 本研究选取稻-麦连作系统(continuous rice-wheat system)和稻-休耕系统(rice-fallow system)各3个样地, 所有样地种植方式已维持连续3年。农田生态系统包括了作物生境和其周围的半自然生境, 如田埂、沟渠等, 因此, 每个样地调查生境包括田内(interior)、田埂(field bund)和沟渠(drainage ditch), 每种生境类型随机选取5个具有代表性的样方进行调查, 其中沟渠的调查范围包括沟渠边坡和沟渠内。根据草本植物调查的样方标准[23], 设定样方面积为(1 m× 1 m或0.5 m× 2 m), 累计样方数量为80个, 其中, 稻-麦连作农田45个样方, 稻-休耕农田35个样方。样方类型分别标记为RWI(稻-麦连作田内)、RWB(稻-麦连作田埂)、RWD(稻-麦连作沟渠)、RFI(稻-休耕连作田内)、RFB(稻-休耕连作田埂)和RFD(稻-休耕连作沟渠)。调查并记录每个样方内的植物种类、盖度、频度, 对于在野外调查期间难以辨认的物种, 要求在原生境下拍照, 并制作标本, 带回室内以作进一步物种鉴定。调查时间为2015年6月。
1.2.2 数据分析 物种多样性测度采用ɑ 多样性的测度方法[24, 25, 26]:物种丰富度指数(R)采用物种数(S); 物种多样性分析采用Shannon-Weiner多样性指数(H); 均匀度分析采用Pielou指数(J); 优势度分析采用Simpson指数(D); 用Jaccard相似系数(Cj)计算稻-麦连作系统和稻-休耕系统的物种相似性。相关参数计算公式如下:
物种丰富度指数:
R=S;
Shannon-Weiner多样性指数:
H'=-lgN
Pielou均匀度指数:
J=-
Simpson指数:
D=1-
重要值:I=(种i相对频度+种i相对盖度)/2;
Cj=c/(a+b-c).
式中:S为每一样地中的植物物种总数目, N为所有物种总个体数, Pi为种i的相对重要值; 相对频度=(某物种频度/总物种频度之和)× 100%; 相对盖度=(某物种盖度/总物种盖度之和)× 100%; a和b分别为两个不同系统的植物种数; c为两个不同系统共有的植物种数。以3个样地的所有样方为基础数据, 根据上述公式分别计算稻-麦连作和稻-休耕系统的各多样性参数; 以各样地不同生境的样方为基础数据, 根据上述公式分别计算稻-麦连作和稻-休耕系统各生境的多样性参数。
排序分析是研究植物群落生态关系的重要方法, 可以揭示植物群落结构及其与环境间的生态关系。只使用物种组成数据时采用非限制性排序方法, 如去趋势对应分析(detrended correspondence analysis, DCA)[27]。本研究采用该法构建群落-物种矩阵, 对6个样地、80个样方的植物群落进行排序分析, 物种在每个样方中的有或无以0和1表示。
数据初步分析、物种多样性计算及方差分析分别在Excel和DPS 13.5软件中完成, 80个样方的群落排序使用Canoco 5.0软件完成。
2 结果与分析
2.1 植物的物种组成分析
稻-麦连作系统的3个样地总计64种植物, 分属于28科58属, 种类较多的有禾本科(16属16种)和菊科(6属9种), 共占物种总数的39.1%; 其次是莎草科(2属4种)。不同生境中, 物种数量和种类差异较大, 其中田埂植物由22科44属49种组成, 占稻-麦连作农田植物物种总数的76.6%; 沟渠植物有16科29属32种; 田内植物共有11科18属20种, 且许多田埂上出现的植物未在田内发现, 如车前(Plantago asiatica)、一年蓬(Erigeron annuus)、狗尾草(Setaria viridis)等。
稻-休耕系统3个样地总计76种植物, 分属于32科68属, 种类较多的科依次为禾本科(14属14种)、菊科(7属11种)、苋科(4属4种)和莎草科(2属4种)。3种生境下的物种数量多少依次为田埂> 田内> 沟渠, 其中田埂植物隶属于29科60属65种, 田内植物共有17科33属37种, 沟渠植物隶属于19科30属33种。与稻-麦连作系统相比, 稻-休耕系统出现许多新的植物种类, 如茅莓(Rubus parvifolius)等蔷薇科植物、马松子(Melochia corchorifolia)等梧桐科植物; 稻田休耕后, 豆科植物小巢菜(Vicia hirsuta)和菊科植物一年蓬(Erigeron annuus)、蒲公英(Taraxacum mongolicum)等数量上升, 而稗(Echinochloa crusgalli)、野老鹳草(Geranium wilfordii)、喜旱莲子草(Alternanthera philoxeroides)等植物数量下降(表1)。比较稻-麦连作和稻-休耕系统的物种相似性, Jaccard系数值为0.49。
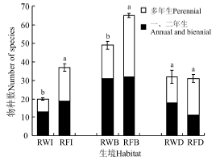
从生活周期角度, 稻-麦连作系统的一、二年生植物有39种, 占物种总数的60.9%; 而稻-休耕系统的一、二年生植物也有36种, 多年生植物有40种, 一、二年生植物占物种总数的47.4%。此外, 稻-麦连作系统45个样方的植物均为草本植物, 稻-休耕系统30个样方植物仍以草本为主, 但是21.7%的样方中出现乔、灌木。此外, 对稻-麦连作和稻-休耕系统相同生境下物种数进行比较, 稻-休耕系统的田内和田埂物种数量显著高于稻-麦连作系统, 但是两个系统的沟渠物种数量无显著差异(图1)。
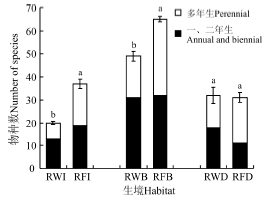 | 图1 稻-麦连作和稻-休耕系统不同生境下的植物物种数注:不同小写字母表示同种生境下不同系统植物物种数差异显著(P< 0.05)。Fig. 1 Number of plant species under different habitats in continuous rice-wheat and rice-fallow systemsNote: Different lowercase letters indicate significant difference in the same habitat type among different systems at the 0.05 level. |
稻-麦连作系统不同生境下的植物物种丰富度差异较大:田内植物有20种, 平均值为13.0; 田埂总计有49种植物, 各样方有植物21~30种, 平均值为26.3; 沟渠总计有32种植物, 各样方平均值为21.0。稻-休耕系统3种生境下的物种丰富度大小依次为田埂> 田内> 沟渠, 其中田埂植物总计65种, 各样方平均值为35.3; 田内植物总计37种, 各样方平均值为21.7, 沟渠植物总计33种, 各样方平均值为25.0(图1, 表1)。
| 表1 稻-麦连作和稻-休耕系统3种生境的植物物种统计 Table 1 Tatistics of plant species in three habitats of continuous rice-wheat and rice-fallow systems |
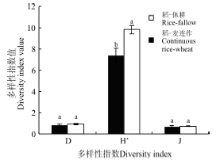
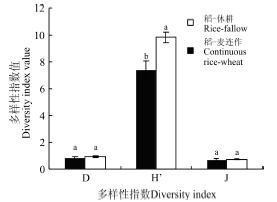
2.2 植物物种的丰富度、多样性和均匀度 稻-休耕系统的物种丰富度指数(R)和物种多样性指数(H')均显著高于稻-麦连作系统(P=0.021; P=0.006), 但是两者优势度指数(D)(P=0.318)和均匀度指数(J)(P=0.844)均无显著差异(图2)。 不同生境下, 稻-麦连作系统Simpson优势度指数、Shannon-Weiner多样性指数差异大, 其中田埂最高, 其次为沟渠, 田内多样性最低。在稻-休耕系统中, 除Pielou均匀度指数外, 田埂各多样性指数(R、H、D)均高于田内(表2)。
2.2 植物物种的丰富度、多样性和均匀度 稻-休耕系统的物种丰富度指数(R)和物种多样性指数(H')均显著高于稻-麦连作系统(P=0.021; P=0.006), 但是两者优势度指数(D)(P=0.318)和均匀度指数(J)(P=0.844)均无显著差异(图2)。 不同生境下, 稻-麦连作系统Simpson优势度指数、Shannon-Weiner多样性指数差异大, 其中田埂最高, 其次为沟渠, 田内多样性最低。在稻-休耕系统中, 除Pielou均匀度指数外, 田埂各多样性指数(R、H、D)均高于田内(表2)。 稻-麦连作和稻-休耕系统不同生境多样性特征 Table 2 Diversity indices of different habitats in continuous rice-wheat and rice-fallow systems | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
相同生境下, 稻-麦连作系统的物种丰富度、Simpson优势度指数和Shannon-Weiner多样性指数均低于同生境下的稻-休耕系统。从Simpson指数来看, 田内的稻-麦连作和稻-休耕D值差异极显著(P=0.006), 田埂(P=0.263)和沟渠(P=0.259)未表现出无显著差异; Shannon-Weiner指数, 田内稻-麦连作极显著低于稻-休耕(P=0.001), 田埂之间差异显著(P=0.011), 沟渠之间无显著差异(P=0.291); Pielou均匀度指数, 田内稻-麦连作显著低于稻-休耕(P=0.016)、田埂(P=0.259)和沟渠(P=0.291)未表现出无显著差异(表2)。
2.3 植物群落DCA分析
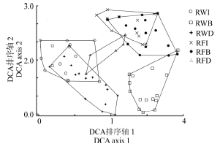
对稻-麦连作和稻-休耕系统不同生境共80个样方进行DCA排序, 得到前4个排序轴的特征值分别为0.501 3、0.407 3、0.299 8和0.270 4, 其中, 第1轴特征值最大, 第2轴次之, 说明前两个排序轴包含更多的生态信息。根据前两个排序轴作二维排序图(图3), 第1轴和第2轴较好地反映了植物群落之间以及群落与环境之间的相互关系。样方DCA图第1轴基本反映了种植制度的变化对群落的影响, 第2轴基本反映了群落沿不同生境类型的变化梯度:对于稻-麦连作系统, 从左到右, 依次为田内、沟渠和田埂; 对于稻-休耕系统, 从左到右, 依次为沟渠、田内和田埂。DCA图中各个样点间距离表示样方内群落相似程度。距离越近, 则说明物种组成越相似; 若出现重叠, 则说明物种组成极为相似。结果表明, 6种类型样地基本相互分离, 但仍有部分重叠, 如RFI和RFB, 说明稻-休耕系统中, 田内和田埂群落组成相似程度高; 相反, RWI和RWB完全分离, 说明稻-麦连作的田内和田埂物种组成差异大。此外, 在稻-麦连作系统中, 除了田埂(RWB)多样性较高外, 沟渠(RWD)和田内(RWI)多样性均较低, 位于图左下方; 相反, 在稻-休耕系统中, 田内(RFI)和田埂(RFB)均具有较高的多样性, 位于图右上方。
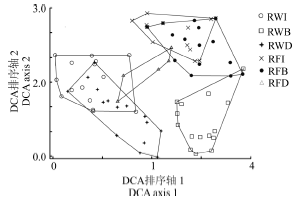 | 图3 80个植物样方的DCA二维排序图Fig. 3 Two-dimensional DCA ordination diagram of 80 quadrats of plant community |
3 讨论与结论
长期稻-麦连作和稻-休耕处理下, 农田植物群落物种组成、相对丰度和盖度等均发生变化, 导致物种丰富度、Shannon-Weiner多样性指数、Simpson优势度指数和Pielou均匀度指数也发生变化, 群落相似性较低(Jaccard系数为0.49)。其原因可能是作物期和休耕期的植物生存的农田环境条件不同, 使不同种类植物的生长发育受到不同程度的影响, 表现为植物群落组成和物种多样性不同。本研究发现, 稻-休耕系统的物种丰富度、Shannon-Weiner指数和Simpson指数均高于稻-麦连作系统, 表明休耕期对植物物种丰富度和多样性有积极作用, 这可能主要源于休耕期停止了化学品的投入和土壤扰动, 改善了土壤营养。
休耕对农田群落的影响首先与农业化学品投入减少有关。研究表明, 不施肥可以提高农田杂草多样性, 这是杂草长期适应土壤条件及与作物竞争的结果[5, 7]。有研究发现, 在停止使用除草剂和化肥后, 植物物种平均数目提高了50%左右, 且除草剂对物种多样性和物种组成的影响远大于化肥的使用[28]。本研究中, 稻田休耕后, 与水稻伴生的田间杂草, 如稗、千金子(Leptochloa chinensis)、异型莎草(Cyperus difformis)和碎米莎草(Cyperus iria)大幅度减少, 而菊科植物如一年蓬、鼠麴草数量剧增, 其它野生植物, 如蒲公英、毛茛(Ranunculus japonicus)和车前(Plantago asiatica)数量也增加。同时, 适氮植物, 如猪殃殃(Galium spurium)数量激增, 而需氮水平高的物种则下降, 为豆科植物所取代, 如小巢菜和天蓝苜蓿。该结果表明, 休耕后土壤养分的变化会直接影响物种组成和相对丰度。
休耕对农田植物群落的影响与土壤扰动的停止有关, 其效应类似于免耕对农田植物群落的影响。有研究表明, 土壤扰动少的免耕系统, 有利于多年生杂草的发生, 因为多年生杂草不会因为频繁的耕作活动而折断死亡, 种子也更易于在土壤表层积累并繁殖[12, 13, 29]。
另有研究发现, 休耕年间与作物种植年间的植物群落组成有较大的不同, 且植物物种多样性有较大提高, 在于休耕期允许土壤种子库的维持, 并可能缓冲了作物期的负面效应[3]。有研究也表明, 土壤扰动少非常有利于更大和更多样化的土壤种子库的建立, 促进植物多样性的提高[16, 30]。相反, 稻-麦连作系统中, 农田长期处于干湿交替和土壤受扰动的状态, 这对种子在土层中的贮存不利[5]。本研究中, 休耕可以促进原来稻田生态系统中由草本群落逐步向草-灌-乔复合群落演替。在连作系统中, 与水稻伴生的主要是一、二年生短生活周期植物, 如繁缕(Stellaria media)和婆婆纳(Veronica polita); 在休耕系统中, 多年生植物增加, 间接反映在停止施用化肥和人工干扰变弱后, 一年生杂草得到了控制。
此外, 无论稻-麦连作还是稻-休耕系统, 物种丰富度和多样性的差异主要存在于田内、田埂和沟渠不同生境之间, 其中田埂各参数值最高。两个系统中, 田埂植物种类占农田生态系统植物物种总数分别为76.6%和85.5%, 且田埂和沟渠的植物物种总数比例可达100%, 表明非作物生境在生物多样性维持上具有重要意义。DCA排序图显示, 稻-麦连作样地和稻-休耕样地几乎完全分离, 说明休耕影响了植物群落组成。且稻-麦连作系统中, RWD和RWI部分重叠, 但与RWB完全分离; 而稻-休耕系统中, RFI和RFB相互重叠, 说明连作促进农田不同生境的植物群落差异, 而休耕可以降低作物生境和田埂的植物组成差异。休耕可以促进植物群落向农田非作物生境(如, 田埂及其它农田边界)群落靠近, 暗示着休耕具有形成农田野生植物避难所的作用[3, 31]。
综上, 稻-麦连作的物种丰富度和各生物多样性指数较稻-休耕系统低。因此, 休耕可有效提高植物多样性, 缓冲了作物期的负面效应。休耕期内, 通过保持土壤种子库和周围自然或半自然生境的种子扩散, 可有效保护植物物种多样性。研究同时表明, 稻-麦连作和稻-休耕系统植物群落组成差异较大, 且休耕能促进多年生植物的发展。本研究中, 休耕导致的植物群落的变化则可能与休耕期农业化学品停止施用、土壤扰动减少以及土壤种子库的保持和周围非作物生境的种子有效扩散有关。
(责任编辑 张瑾)
The authors have declared that no competing interests exist.
参考文献
| [1] |
|
| [2] |
|
| [3] |
|
| [4] |
|
| [5] |
|
| [6] |
|
| [7] |
|
| [8] |
|
| [9] |
|
| [10] |
|
| [11] |
|
| [12] |
|
| [13] |
|
| [14] |
|
| [15] |
|
| [16] |
|
| [17] |
|
| [18] |
|
| [19] |
|
| [20] |
|
| [21] |
|
| [22] |
|
| [23] |
|
| [24] |
|
| [25] |
|
| [26] |
|
| [27] |
|
| [28] |
|
| [29] |
|
| [30] |
|
| [31] |
|