

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
城市绿地生态系统碳交换动态及其与环境控制因子的关系
引用本文
王亚军, 郁珊珊. 城市绿地生态系统碳交换动态及其与环境控制因子的关系. 草业科学, 2017,34(5):966-974
Wang Ya-jun, Yu Shan-shan. Dynamics of CO2 exchange and its environmental controls in an urban green-land ecosystem in Fuzhou City . Pratacultural Science,2017,34(5): 966-974
Permissions
Wang Ya-jun, Yu Shan-shan. Dynamics of CO2 exchange and its environmental controls in an urban green-land ecosystem in Fuzhou City . Pratacultural Science,2017,34(5): 966-974
Copyright©2017, 《草业科学》编辑部
《草业科学》编辑部
城市绿地生态系统碳交换动态及其与环境控制因子的关系
摘要
随着城市的不断发展,城市绿地面积逐年增加,城市绿地生态系统潜在的碳汇和碳源功能显得非常重要。2014年1月至2016年1月,基于涡度技术对福州市绿地生态系统进行了碳通量连续两年的定位观测,并探讨了城市绿地生态系统碳交换年际动态特征及其环境调控因子。结果表明,城市绿地生态系统年总生态系统生产力(GEP)、生态系统净生产力(NEP)和生态系统呼吸(Re)在年际之间呈现出一致的变化规律,其中生态系统碳交换特征表现为生长季(4月–11月)以吸收CO2为主,非生长季以释放CO2为主。城市绿地Re均与GPP、GEP呈正比关系,并且拟合线性斜率<1,受生长季的降水量影响较大。采用指数方程对碳交换和土壤温度(Ta)进行模拟,生态系统日平均净碳交换量(NEEday)随Ta的增加呈指数增长模型,Ta可解释80%以上的变异;GEP随着Ta的增加逐渐增加,当Ta达到最高时,并未对GEP产生抑制作用;当气温低于5 ℃时,城市绿地生态系统以呼吸作用为主导,当气温高于5 ℃,城市绿地生态系统净吸收大气CO2为主导。光合有效辐射(PAR)与NEE呈直角双曲线关系,当光补偿点PAR小于350 μmol·(m2·d)–1时,NEE为正值,此时生态系统呼吸大于光合生产;当PAR大于500 μmol·(m2·d)–1时,生态系统呼吸与光合生产持平,其碳吸收量基本达到饱和。生态系统最大光合速率、白天平均生态系统呼吸强度、表观量子效率(α)与温度和PAR的季节变化趋势相一致,均在7月最大和5月最小。相关性分析可知,不同月份NEE残差均与降水量呈负相关关系,与饱和水汽压差(VPD)呈正相关关系。上述研究结果为估算、模拟和预测城市绿地生态系统碳交换提供了基础数据支持和理论基础。
关键词:
涡度技术; 城市绿地生态系统; 生态系统生产力; 生态系统净生产力; 生态系统呼吸
中图分类号:S181
文献标志码:A
文章编号:1001-0629(2017)05-0966-09
doi: 10.11829/j.issn.1001-0629.2016-0435
Dynamics of CO2 exchange and its environmental controls in an urban green-land ecosystem in Fuzhou City
Abstract
: The area of urban green-land is expanding dramatically as a strategy to counter rapid urbanization. Urban green-land ecosystems with plantations as their main vegetation type have great potential to sequester atmospheric carbon. Continuous measurements of CO2 flux were made using eddy covariance technique, from January 2014 to January 2016, in Fuzhou City to quantify the seasonal dynamics of net ecosystem CO2 exchange (NEE) and its responses to environmental factors. Gross ecosystem productivity (GEP), ecosystem respiration (Re), and net ecosystem productivity (NEP=–NEE) showed strong seasonal pattern, with CO2 uptake dominating during the growing season from April to November, and a respiratory release of CO2 dominating during the non-growing season. Ecosystem respiration (Re) had a positive relation with GEP and gross primary productivity, and the fitting line was less than 1, with the straight intercept not zero, which shows that the balance of urban green-land ecosystem was greatly influenced by the precipitation and distribution of precipitation during the growing season. An exponential equation of the net carbon exchange to simulate the soil temperature (Ta) showed that NEE increased exponentially with the Ta, and the interpretation ratio was more than 80%. GEP increased exponentially with the Ta, and there was no optimum temperature, i.e., the highest temperature had no inhibitory effect on the GEP. NEP decreased with increasing Ta, when Ta <5.0 ℃, but increased when Ta <5.0 ℃. NEE and photosynthetically active radiation (PAR) meet the optimal hyperbolic relationship when PAR <350 μmol·(m2·d)–1, where the ecological system breathing was greater than the photosynthetic production, and when PAR > 500 μmol·(m2·d)–1, where the ecosystem carbon uptake was saturated. The ecosystem quantum yield (α) and maximum photosynthesis (Amax) showed apparent seasonal patterns, both peaking in July. Correlation analysis showed that the NEE residuals for different months were negatively correlated with rainfall and positively correlated with Ta and vapour pressure deficit. The present results could contribute to the carbon budget of urban ecosystems and help create carbon-oriented management strategies for sustainable urban development under global climate change.
Keyword:
eddy covariance technique; urban green-land ecosystem; gross ecosystem productivity; net ecosystem productivity; ecosystem respiration
城市是一类以人类活动为中心的社会、经济和自然复合生态系统。城市绿地是城市中自然生态系统的基础部分, 以植被为主体, 以土壤为基质, 以自然和人为因素干扰为特征, 在生物和非生物因子协同作用下所形成的有序整体[1]。城市绿地系统是城市生态系统的重要组成部分, 通过系统结构来实现系统中物质循环和能量转化过程[2]。随着城市生态概念的发展, 城市绿地生态系统具有重要的意义和价值, 然而工业化给城市绿地生态系统环境造成了潜在的威胁, 并且加速了城市生态环境危机[2]。受人类活动的影响, 城市绿地生态系统表现出高CO2浓度、高温、高气溶胶等特征[3]。气体悬浮物极大削弱了城市地面对太阳辐射的接受, 然而却增加了散射辐射的比例; 同时, 大气悬浮气溶胶粒子增加了低层大气中上部温度, 从而造成单位面积碳固持能力增加或降低[4, 5]。作为改善和美化城市生态环境的重要载体, 城市绿地生态系统是各类草地、耕地、林地、公园等系统的总称, 具有一定的自净能力、自动调节能力和生命力, 是城市生态环境净化的“ 调节器” , 在调节城市生态环境平衡中发挥着重要作用[6]。与此同时, 在碳循环与气候变化研究中, 绿地生态系统对减轻气候变化的影响发挥着重要作用, 其潜在的碳汇功能(或碳固持)也不可低估[7, 8]。然而, 目前相关的研究仅仅表现在有关城市热岛和城市污染等碳源方面的报道, 关于城市绿地生态系统碳汇功能对城市生态系统重要性的研究仍相对薄弱[6, 7, 8]。通过对城市绿地生态系统碳汇功能的量化研究, 可以评估并预测其在减少大气CO2中的潜在作用, 对城市绿地生态系统碳交换规律及其环境控制因素的理解和认识对我国陆地生态系统碳循环和气候变化研究具有重要的理论和实践意义。
随着全球变化与大气增温(主要表现为CO2浓度的富集), 减缓和适应全球气候变化已成为当前生态学领域研究的焦点之一[9, 10]。早在1996年, 欧美科学家利用涡度技术初步建立了全球CO2通量网, 进而在全球范围内开展了不同尺度生态系统碳交换特征, 但绝大部分研究是针对森林类型, 对草地生态系统碳交换的研究也主要集中在低海拔的平原地区[9, 10]。自20世纪90年代, 我国应用涡度相关法开展了陆地生态系统碳交换特征, 尽管起步较晚, 但也取得了部分有价值的研究进展, 其成果主要集中在森林和草地生态系统碳交换及其对环境的响应等方面[11, 12, 13]。相比之下, 城市绿地作为镶嵌在城市生态系统的不连续体, 人工管理、热岛效应等人为管理措施更增加了碳交换的不确定性及可变性, 有学者认为, 可以将城市绿地生态系统作为研究生态系统对全球气候变化的响应和预测未来生态系统的变化趋势的“ 自然大实验场” [2, 3, 7], 通过对城市绿地生态系统的碳汇和碳源的连续定量研究, 能够预测其城市生态系统碳交换与收支。目前, 最为广泛的研究是采用涡度相关法测定大气与土壤碳交换, 其测定的空间范围能够延续数十平方米, 也能够达到连续动态监测的目的。这使得对我国陆地生态系统碳交换的长期定位观测成为可能, 如今已广泛应用于陆地生态系统监测中[14]。涡度相关系统可以对外界温度、风速、CO2脉冲值及其大气C交换连续监测, 也有助于理解城市绿地生态系统与CO2交换规律及其动态特征[15, 16, 17]。有鉴于此, 本研究采用涡度协方差法于2014年1月至2016年1月对福州市绿地生态系统进行连续监测, 并结合碳通量的定量数据, 分析城市绿地生态系统净生产力(NEP)、生态系统总生产(GEP)、生态系统呼吸(Re)的季节动态特征以及环境调控因子, 旨在探索城市绿地生态系统对气候变化的响应机制, 为城市生态系统对环境变化的响应及碳循环提供数据支持和科学依据。
1 材料与方法
1.1 研究区概况
福州市位于福建省东部、闽江的下游(118° 08′ – 120° 31′ E, 25° 15′ – 26° 39′ N), 该区气候温暖湿润, 资源丰富, 霜少无雪, 雨量充沛, 农业活动较大, 地貌类型以丘陵为主, 地势起伏不平, 地形复杂多样, 土壤类型较多, 主要包括红壤、黄棕壤、紫色土、潮土和沼泽土, 平均海拔9.2 m, 年均降水量为900~2 100 mm, 年均日照时数为1 700~2 000 h, 无霜期达329 d, 年均气温为16~20 ℃, 年相对湿度约为76%。福州市土地总面积为12 251 km2, 截止到2015年, 绿地面积约为81.2 km2, 人均绿地面积约11.2 m2。
1.2 测定方法
在绿地斑块方面, 福州市绿地存在分布格局呈现点、块状分布, 城区缺少足够宽的绿地廊道进行连接, 公共绿地分布不均, 公园绿地的占地面积最大。由此, 根据福州市城市绿地分布格局及特点, 将涡度通量观测仪安装在福州市西湖公园, 该绿地生态系统保持较为完整, 能够代表城市绿地生态系统, 位于福州市区中心区域, 至今有1 700多年的历史, 是福州保留最完整的一座古典园林, 占地面积42.51 hm2。安装高度约12 m, 包括三维超声风速仪、碳水(CO2/H2O)红外气体分析仪。光量子传感器和净辐射仪安装高度均为12 m, 与涡度通量观测仪安装高度保持一致, 在距离通量塔3 m和6 m迎风位置安装两个翻斗式量雨器, 分别在2、5、8、10和12 m高度上各安装5个空气温、湿度传感器, 然后在塔周边10 m的范围内沿5个方向分别均匀地布设5个土壤水分和土壤温度传感器, 安装深度为10 cm。最后以10 Hz的频率记录和搜集涡度协方差系统的数据, 微气象系统记录每30 min的平均值。
数据采集和质检每周进行一次, 每个月用露点发生器和高纯氮气分别对H2O和CO2零点标定, 其中供电系统为220 V交流电, 蓄电池作为主要的备用电源。观测项目主要包括气象要素观测场、近地面层涡动相关湍流、土壤温度和湿度、自然条件下的风速和风向, 其中辐射观测包括长波辐射和太阳辐射。
1.3 涡动相关资料数据处理
本研究所用数据从2014年1月至2016年1月, 期间由于仪器故障、停电(自然因素)、人为干扰等影响造成了大约10%涡度数据遗失, 因此, 采用EdiRe软件对数据进行校正, 每个数据样本处理时间均为0.5 h, 处理步骤如下:1)利用3倍标准差法剔除由停电、仪器故障、人为干扰等造成的异常值和明显存在错误的野点; 2)将平均量、方差、脉动量、协方差等进行二维坐标轴旋转; 3)通过密度脉动订正(WPL)和高频损失订正等对碳通量进行修正; 4)物理检验和湍流平稳性检验。通过以上所有通量资料和数据, 按照Lindroth等[18]、Winter和Schramm[19]进行数据修订与质量控制。
城市绿地生态系统净碳交换(NEE)计算公式为[20]:
NEE=Fc+Fs+Vc.
式中:Fc代表通量观测塔所测的CO2通量, Vc表示水平和垂直的平流效应, Fs代表涡度相关系统冠层内储存通量。地势平坦、植被分布均匀区域的Vc可忽略不计。对于较高的冠层, Fs一般并不为零, 因此, 需要对Fs进行矫正和计算[20];
Fs=
式中:Δ CZ表示观测高度处的CO2浓度(μ mol· m– 3), Δ Z表示观测高度(m), Δ t表示采集数据周期(s)。对夜间NEE数据(NEEnight)进行摩擦风速的筛选, 剔除风速< 0.15 m· s– 1的数据。通过足迹分析, 认为上风向为90%通量足迹(范围为150 m), 100 m的贡献率为70%, 而通量贡献的峰值则出现在离地面高度大概50 m处。此外, 对于白天光合有效辐射(PAR)高于3 μ mol· (m2· d)– 1的数据采用如下光响应模型进行插补[20]:
NEEday=
式中:NEEday表示白天的生态系统净碳交换量[μ mol· (m2· d)– 1], α 表示生态系统表观光量子效率(μ mol· μ mol– 1), PAR表示光合有效辐射, Amax表示生态系统最大光合速率[μ mol· (m2· d)– 1], Rd表示白天平均生态系统呼吸速率[μ mol· (m2· d)– 1]。
生态系统呼吸(Re)与土壤温度常用的关系模型有以下4种。
Taylor和Lloyd模型[21]:
Re=R10
Arrhenius模型[21]:
Re=R10
指数模型[22]:
Re=R10
Q10模型[22]:
Re=R10
式中:Ea表示活化能(温度敏感系数); R10表示10 ℃的生态系统呼吸速率, R表示气体常数E0=Ea/R; Tref表示参考温度(10 ℃); T表示土壤温度或气温; Ts表示土壤温度; T0表示绿地生态系统呼吸=0的温度; Q10表示温度的敏感性指标, 也即温度每改变10 ℃, 呼吸速率涉及的改变范围, 本研究在Taylor和Lloyd模型[23]的基础上进行改进, 从而Q10模型也可表示为Re=ln(Q10/10)。
生态系统呼吸(Re)与土壤水分常用的关系模型有以下2种。
连乘模型[23]:
Re=a× eb× T×
线性模型[23]:
Re=a× vwc+b .
式中:vwc为土壤体积含水量, a, b, c为待拟合参数。
本研究分别采用线性方程对夜间Re与土壤温度关系进行拟合, 为了综合土壤水分、温度对Re的影响, 利用连乘模型来估算白天Re。
Re=Reday+Renight .
式中:Reday为白天生态系统呼吸, Renight为夜间生态系统呼吸。
GEE=NEE– Re .
总生态系统生产力(GEP)与GEE的绝对值在数值上相等并且符号相反。因此, 从生态系统的角度上考虑, 可以认为生态系统总初级生产力(GPP)与GEP相等, 也即GEP=– GEE=GPP。
数据统计分析与作图均采用SPSS 21和Origin 9.2完成, 通过回归方程及相关性分析研究生态系统碳交换及其组分与环境因子之间的关系。
2 结果
2.1 微气象因子的季节变化
本研究城市绿地生态系统最高气温(Ta)出现在7月, 最低气温出现在1月, 3月– 11月平均气温均为正值(图1)。PAR呈单峰型的变化趋势, 最大值出现在6月, 其平均值为365.24 μ mol· (m2· d)– 1, 最小日总量出现在1月, 平均为132.54 mol· (m2· d)– 1, 同时受天气影响(如雨雪和沙尘), PAR存在较大的短期波动。年降水总量为1 124.56 mm, 最大日降水量达到59.23 mm, 80%以上降水集中在6月– 9月, 其中7月降水量最高(198 mm)。饱和水汽压差(VPD)在0.16~2.79 kPa, 呈现出明显的季节变化规律, 总体表现为夏季和秋季较高, 春季和冬季较低。
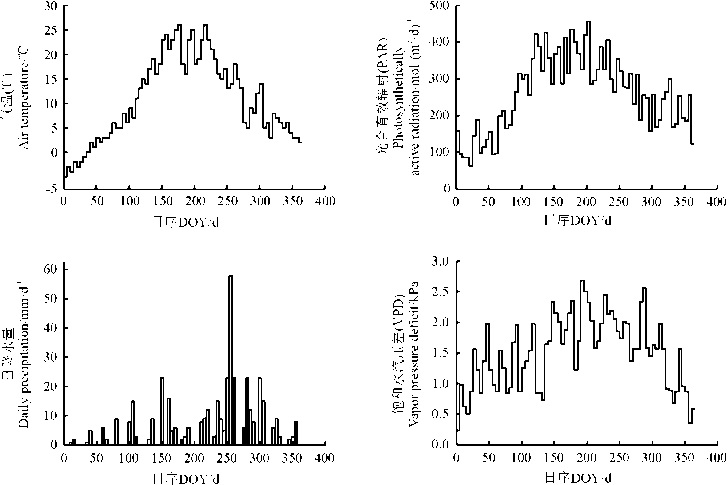 | 图1 微气象因子的季节变化 |
2.2 生态系统净碳交换的季节变化
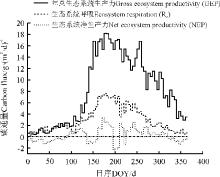
城市绿地生态系统GEP、NEP和Re年总量分别为4.19× 105、0.49× 105、3.74× 105 g C· m– 2(图2)。6月– 8月为明显的碳吸收阶段, 生长季中期无降水时, 土壤水分起到了一定的胁迫作用, 因此城市绿地生态系统植被光合作用在7月以后急剧降低, 其中日最大GEP约18 g· (m2· d)– 1。本研究中, 3、4月份绿地生态系统植被萌芽较早, 导致3月份绿地生态系统GEP高于Re, 该时期表现为净碳吸收时期; 而5、6月份降水较少, NEP出现降低的阶段, 此后, 随着雨季的来临, 城市绿地系统表现出明显的净碳吸收, NEP逐渐增大; 在7、8月的中旬最大, 并且一直持续到9月, 生态系统净碳吸收作用才开始有所降低。
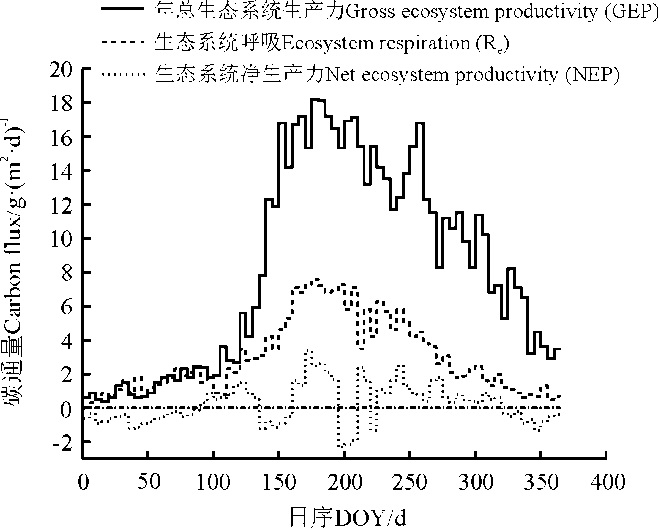 | 图2 生态系统净碳交换的季节变化 |
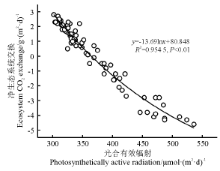
2.3 碳呼吸对生态系统碳交换的影响
城市绿地Re与GEP、NEP具有一定的相关性(图3)。城市绿地Re与GEP显著正相关(P< 0.01), 同时二者的直线截距均不等于0, 拟合直线斜率均小于1。城市绿地生态系统dRe/dGPP均为正值, Re随GPP增加呈一定的增加趋势, 表明福州市绿地生态系统受到环境因子年际波动的影响较为显著, 导致该生态系统结构和功能发生了一定的变化。另外, 福州市年降水较少, 生态系统植被生长季较晚和异养呼吸值较高, 尽管在7、8月份雨季来临以后生态系统固碳功能逐渐恢复, 但整个生长季植被固碳均为较弱的碳源。从总体上来看, 城市绿地生态系统Re与NEP和GEP比值均< 1, 并且净碳交换年际变异较大, 主要受环境因素的影响(降水), 该系统的碳汇和碳源功能会发生一定的转变。由此可知, 受降水的影响, 城市绿地生态系统可以分别为弱的碳源和较强的碳汇, 并且净碳吸收功能与净碳吸收持续时间也存在较为密切的相关关系。
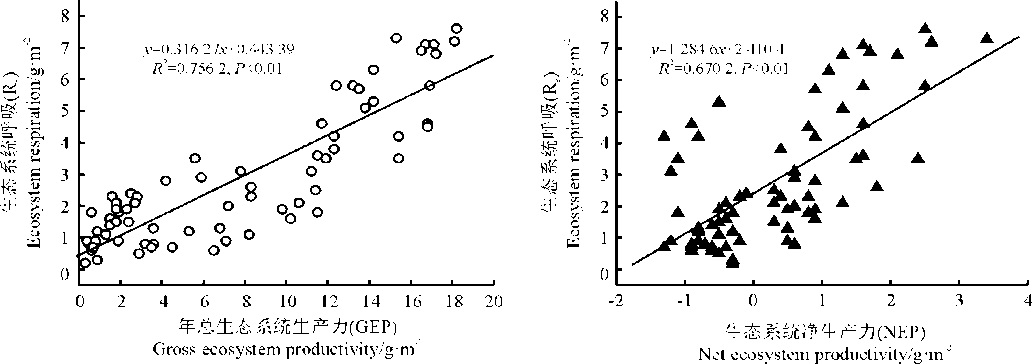 | 图3 碳呼吸对生态系统碳交换的影响 |
2.4 温度对净生态系统碳交换的影响
通过SPSS最佳拟合度筛选分析后, 运用指数方程拟合净碳交换和土壤温度之间的关系, 其中10 cm土壤温度可以很好地解释生态系统的呼吸变异性, 经计算可知, 城市绿地生态系统呼吸的温度敏感性(Q10)的值为2.2 ℃(图4)。生态系统日平均NEE随着Ta呈指数增长, 并且Ta能够解释80%以上NEE的变异。温度不仅可以影响绿地生态系统的呼吸, 还可以影响到生态系统碳交换的形式。本研究中, 春季GEP随着Ta的升高呈逐渐增加趋势, 夏季达到最高, 秋季Ta有所降低, 导致GEP也开始降低。当气温低于5 ℃时, 该生态系统以呼吸作用占主导作用, 此时生态系统地上植被处于非生长季, Re随温度的增加而增加, 导致日总NEE随温度呈增加的趋势。当气温高于5 ℃时, 该生态系统植被进入生长季, 地上植被的光合作用增加的幅度较呼吸作用快, 表现为净吸收大气CO2, 其中5 ℃时生态系统表现为最大吸收, 之后随着气温的继续增加, 日总NEE逐渐降低, 但总体来看, 日总NEE与Ta呈指数曲线关系。
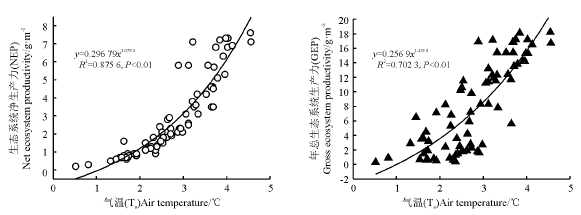 | 图4 温度对净生态系统碳交换的影响 |
2.5 光合有效辐射与净生态系统交换的关系
NEE与PAR之间呈显著的直角双曲线关系, 在光补偿点PAR< 350 μ mol· (m2· d)– 1时, NEE为正, 此时生态系统光合作用小于呼吸作用; 随着光补偿点PAR的增加, 生态系统光合作用和净碳吸收逐渐增加; 当PAR> 500 μ mol· (m2· d)– 1时, 生态系统的碳吸收量达到饱和状态(图5)。表观量子效率变化范围在0.021 6~0.056 9 μ mol CO2· μ mol– 1, 其中白天平均生态系统呼吸强度和生态系统最大光合速率与PAR、温度变化趋势相一致(7月最大, 5月最小)(表1)。光合作用随着PAR呈逐渐增强趋势, NEE的值也随之增加, 生态系统的碳汇能力也逐渐增加, 其中平均白天生态系统呼吸速率和最大光合速率和均在7月最大, 与光合有效辐射季节变化趋势相一致。
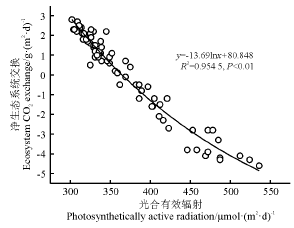 | 图5 光合有效辐射与净生态系统交换的关系 |
| 表1 生长季模拟的光合参数 Table 1 Photosynthetic parameters modelled in the growing season |
2.6 环境因子对NEE的影响
为了合理解释NEE与土壤温度(Ta)、日降水量和饱和水汽压差之间(VPD)的相关性, 利用NEE残差与土壤温度(Ta)、日降水量与饱和水汽压差(VPD)分别做Pearson相关性分析, 结果表明, 不同月份NEE残差均与日降水量负相关, 其中7、8月份的NEE残差与日降水量呈极显著相关(P< 0.01)(表2); 不同月份NEE残差均与Ta和VPD正相关, 其中7、8月份的NEE残差与VPD呈极显著相关(P< 0.01); 5、6、7、8月的NEE残差与Ta极显著正相关(P< 0.01), 9、10月的NEE残差与Ta显著正相关(P< 0.05)。
| 表2 环境因子与NEE残差的相关性 Table 2 Correlation between NEE residual and main environmental factors |
3 讨论
生态系统的碳循环是全球研究的热点问题之一, 然而城市绿地生态系统碳循环研究则成为陆地碳循环研究进展的瓶颈[11]。对城市绿地生态系统CO2通量的连续定点监测将有助于分析城市绿地生态系统碳收支的动态平衡[11, 12, 13]。本研究监测了城市绿地生态系统2年的通量数据, 与土壤、空气温度、大气太阳辐射等气象数据相结合, 利用涡动相关法分析了城市绿地生态系统碳交换特征及其环境调控因子。结果表明:城市绿地生态系统净碳交换过程具有明显的时空和季节性差异。NEE在7月– 9月最大, 在此过程中, 降水量和降水的时空分布特征对其影响较大。从5月份开始, 绿地生态系统表现出明显的碳汇功能, 而生长季无植被受水分胁迫导致净碳吸收作用受到抑制, 从而净碳吸收过程均呈现“ 双峰” 的变化趋势。有研究认为[20, 21], 陆地生态系统本身是一个具有自身固碳的有效碳库, 而不同生态系统Re/GPP在理论上< 1; 当GEP为0时, 该生态系统的自养呼吸=0; 对于本研究而言, 城市绿地生态系统拟合直线的截距并不等于0, 而且大于0, 其表示的含义是当城市绿地生态系统不存在植被时, 该系统自身固有的异养呼吸值。
一般认为, 温度是控制生态系统呼吸的主要环境调控因子, 然而以大气还是土壤的温度作为判别的环境调控因子仍没有明确的定论[22]。Ma等[23]认为, 土壤温度与空气温度具有密切的相关性, 二者能够共同反映生态系统呼吸变化趋势; 然而, 由于城市绿地生态系统扰动性比较大, 土壤呼吸可能是决定生态系统呼吸的主要因素之一。因此, 本研究以土壤温度作为驱动因子来解释变量显得更为科学和合理。由于涡动系统自身的局限, 白天NEE不能够完全被预测, 而其它环境因子会对NEE的值产生一定的影响[24]; 另一方面, VPD和温度在一定程度上影响植物的固碳作用, VPD与温度之间也有较强的关联性。内蒙古荒漠草原日尺度的NEE受到20– 30 cm的土壤含水量影响最大[25]; 温带森林土壤水分(VPD和VWC)限制植物生长的关键因子, 而水分亏缺是造成植物固碳量低的主要原因[26]; 在温带落叶森林, 植物生长季节受到降水的影响而提前, 其中生长初期和中期的降水对碳通量的影响比生长后期更为明显[27]。也有研究[11, 20, 21]表明, GEP随温度呈抛物线的变化趋势, 当达到其最适温度时, GEP最大(抛物线的顶点), 以后GEP将有所降低, 表明, 当生态系统Ta达到一定的阈值(临界温度)后, Ta对GEP产生了一定的抑制作用。然而, 本研究中当Ta达到最大时, GEP也没有被抑制。
本研究中, 由于城市绿地生态系统地表的扰动比较大, 导致土壤呼吸值偏高。2015年, 城市绿地生态系统土壤呼吸总量为4.49× 104 g· m– 2, 这远小于北京奥林匹克森林公园[28]的1.73× 105 g· m– 2。此外, 由于绿地生态系统植被光合固碳能力有限(叶面积指数小), 其表观量子效率的变化范围在0.021 6~0.056 9 μ mol· μ mol– 1, 略小于温带森林(0.024~0.085 μ mol· μ mol– 1), 而略大于北京奥林匹克森林公园(0.056 9 μ mol· μ mol– 1)。这主要是由于城市中大气悬浮物极大地削弱了地面接收的太阳辐射和植物的光合作用。本研究中, 城市绿地生态系统光合利用效率强度高于太阳直射辐射, 碳收支对太阳直射辐射具有一定的敏感度, 这与前人[11, 20, 21]的研究结果相吻合。城市绿地生态系统在景观布局、植物结构和人为管理、扰动等方面的差异, 导致本研究在区域尺度的空间代表性上依然不足。在未来的研究中, 需要结合多个绿地生态系统监测站点的数据, 并通过模型参数化和数据整合实现生态系统到区域尺度的发展, 从而实现气候变化背景下预测城市绿地生态系统对路对生态系统碳收支的贡献。
(责任编辑 武艳培)
The authors have declared that no competing interests exist.
参考文献
| [1] |
|
| [2] |
|
| [3] |
|
| [4] |
|
| [5] |
|
| [6] |
|
| [7] |
|
| [8] |
|
| [9] |
|
| [10] |
|
| [11] |
|
| [12] |
|
| [13] |
|
| [14] |
|
| [15] |
|
| [16] |
|
| [17] |
|
| [18] |
|
| [19] |
|
| [20] |
|
| [21] |
|
| [22] |
|
| [23] |
|
| [24] |
|
| [25] |
|
| [26] |
|
| [27] |
|
| [28] |
|