





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
不同盐浓度下硅对高羊茅根系特征的影响
[宋锐, 林选栋, 林丽果, 宋浩然, 刘慧霞 ]
]
]
|
|


第一作者:宋锐(1989-),男,山西大同人,在读硕士生,研究方向为饲草料资源与生态修复的研究。E-mail:[email protected]
利用盆栽试验研究了不同盐浓度生境下硅对高羊茅( Festuca arundinacea)根系生物量和根系形态特征的影响。结果表明,在盐浓度0-300 mmol·L-1的范围内,高羊茅根系生物量、根系表面积、根系体积、根系平均直径和根系总长均随盐浓度增加而呈降低趋势,高羊茅根冠比随盐浓度增加呈现先增加后降低趋势。硅对高羊茅根系生物量和根系特征的影响与盐浓度密切相关,当盐浓度≤150 mmol·L-1时,硅显著提高了高羊茅根系生物量、根系表面积和根系总长( P<0.05),说明硅在一定盐浓度下通过增加高羊茅根系生物量,改变根系形态特征,提高高羊茅在盐生境的生长能力。
A pot experiment was conducted to determine the effect of silicon supply on the biomass and the root features of tall fescue ( Festuca arundinacea) seedlings under different salinity conditions. This study showed that the root dry biomass, total surface of roots, root bulk, total length of roots,and average root diameter were decreased, but the root to shoot ratio of tall fescue seedling initially increased and then decreased with an increase in the salt concentration from 0 to 300 mmol·L-1. The contribution of the silicon supply to root biomass and root features was closely related to the salt concentration: the silicon supply significantly increased the root biomass, the total surface of roots, and the total length of roots ( P<0.05) when the salt concentration was ≤150 mmol·L-1. These results suggested that silicon supply could improve the adaptation ability of tall fescue in different salinity conditions by the alteration of the root biomass and root features.
草坪绿地能够净化空气、美化环境, 是现代城市的重要组成部分[1, 2]。根系是草坪草吸收和利用土壤中水分、营养物质和微量元素的重要器官[3], 直接与土壤环境接触, 也是草坪草最先遭受逆境的重要部位[4, 5]。土壤盐浓度较低时, 草坪草可通过调节地上以及根系形态和生理特征, 适应盐生环境; 而当土壤盐浓度过高时, 草坪草则往往遭遇离子胁迫和渗透胁迫, 从而导致生物量下降, 甚至植株干枯, 根系死亡[5, 6, 7]。
水肥管理是提高草坪草耐盐性的重要途径之一[8]。添加氮、磷和硅均能够提高植物的耐盐性[9, 10, 11], 但氮和磷过量使用会引起水体富营养化和土壤非点源污染[12], 因此环境友好型元素硅成为提高植物耐盐性的重要矿质添加元素[1, 13, 14]。研究表明, 添加硅能够提高小麦(Triticum aestivum)[15]、黄瓜(Cucumis sativus)[16]、玉米(Zea mays)[17]、马齿苋(Portulaca oleracea)[8]和番茄(Lycopersicon esculentum)[18]等植物的耐盐性, 从而提高植物在不同盐浓度生境下的生长能力, 其主要原因是添加硅能够改进植物的根系特征, 增加根系生物量, 促进植物在不同盐生环境下的生长。
高羊茅(Festuca arundinacea)是一种常见的冷季型多年生草坪草, 广泛应用于我国草坪绿地的建设[6]。虽然已有研究证明不同浓度盐生境下, 添加硅能够提高高羊茅地上部分渗透调节, 降低盐胁迫对膜结构的伤害, 增强高羊茅耐盐性[6], 但是根系的变化直接影响着地上部分应对逆境的能力, 与地上相比根系对植物更加重要[8]; 不同盐生环境下硅是否通过改变高羊茅根系生物量、根系形态特征来适应盐生环境, 尚需科学试验提供佐证。本研究通过盆栽试验, 研究不同盐生境下添加硅对高羊茅根系生物量、根冠比、根系体积、根系平均直径、根系表面积和根系总长的影响, 以期阐明硅在不同盐生境下对高羊茅根系生物量和特征的响应。
采用盆栽试验, 供试的高羊茅品种为向导, 来源于百绿国际草业有限公司。试验采用盐浓度和硅双因素处理, 其中盐浓度设为0、50、100、150、200、250和300 mmol· L-1, 每个盐浓度下设置加硅(+Si)和不加硅(-Si)两个处理, 共计14个处理, 每个处理6个重复。盐生境下硅浓度为2 mmol· L-1时高羊茅的种子发芽率和保苗率最高, 生长能力最强[19], 因此, 本研究硅处理的硅离子浓度设置为2 mmol· L-1, 硅源采用硅酸钠。盐浓度处理以NaCl为盐离子来源, 计算Na+浓度时包含了硅酸钠所携带的Na+含量。本研究于2015年5月20日在兰州大学智能温室内进行, 采用高15 cm, 盆口直径20 cm的塑料盆, 每个花盆内播种饱满的高羊茅种子300粒, 基质为珍珠岩和蛭石(体积比为1∶ 1)的混合基质, 然后将塑料盆随机摆放。根据试验设计, 在Hoagland营养液中添加相应浓度的硅酸钠和氯化钠, 将其充分混合后配置成处理溶液, 种植用处理溶液按照试验设计浇灌每个塑料盆, 直至盆底托盘有处理溶液渗出为止, 隔日浇灌一次, 待高羊茅品种完全出苗后(高羊茅发芽时间21 d), 每个盆中选择长势均匀的50株幼苗, 作为继续培养的幼苗, 其它多余幼苗以间苗的形式手工除去, 以便排除密度对高羊茅生长的影响。然后按照试验设计隔天浇灌处理溶液, 至61 d时取样。
每个处理采用3个塑料盆内的植株测定生物量, 用去离子水冲洗干净, 用剪刀将地上、根系分开, 将其置于105 ℃的烘箱杀青15 min, 然后在75 ℃下烘干至恒量, 分别称其干质量, 测定根冠比。其余3个盆收集完整植株, 取地下部分, 用去离子水冲洗干净, 采用0.075 mg· mL-1的甲基蓝将根系浸泡染色10 min后, 用Delta-T SCAN根系分析系统(HP.C7717, Singaporean)扫描根系, 测定总表面积、根系体积、平均直径及根系总长。
先采用Two-Way ANOVA 进行双因素方差分析, 若差异显著再采用Duncan分析法进行多重比较, 统计软件为SPSS 19.0。
硅、盐浓度及其互作均极显著影响了高羊茅根系生物量(P< 0.01)(图1)。随盐浓度增加, 高羊茅根系生物量显著降低(P< 0.05), 盐浓度≤ 250 mmol· L-1时, 高羊茅根系生物量随盐浓度增加显著降低。当盐浓度≥ 200 mmol· L-1时, 硅对高羊茅根系生物量没有显著影响; 但当盐浓度≤ 150 mmol· L-1时, 硅添加能显著提高了高羊茅根系生物量(P< 0.05); 盐浓度为150 mmol· L-1时, 添加硅较不添加硅根系生物量增幅最大, 为34.4%; 盐浓度为50 mmol· L-1时, 添加硅较不添加硅根系生物量增幅最小, 为24%。
 | 图1 不同盐浓度下硅对高羊茅根系生物量影响注:不同小写字母表示不同处理间差异显著(P< 0.05)。下同。Fig. 1 Effect of the addition of silicon on the root biomass of tall fescue at different salt concentrationsNote:Different lowercase letters indicate significant difference among different treatment at the 0.05 level; similarly for the following figures. |
硅和盐浓度均极显著影响了高羊茅根冠比(P< 0.01), 但硅与盐浓度互作对其无显著影响(P> 0.05)(图2)。高羊茅根冠比随盐浓度增大呈现出先上升后下降的趋势, 在盐浓度为150 mmol· L-1时达到最大(P< 0.05); 硅显著增加了高羊茅根冠比(P< 0.05)。
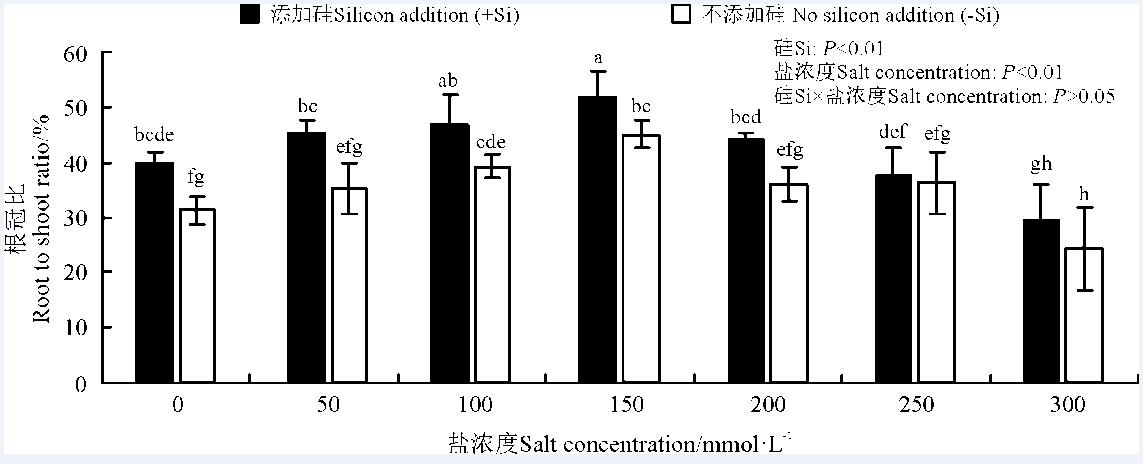 | 图2 不同盐浓度下硅对高羊茅根冠比的影响Fig. 2 Effect of the addition of silicon on the root to shoot ratio of tall fescue at different salt concentrations |
硅、盐浓度及其互作均极显著影响了高羊茅根系总表面积(P< 0.01)(图3)。随着盐浓度增加, 高羊茅根系总表面积显著减少(P< 0.05)。盐浓度≤ 200 mmol· L-1时, 高羊茅根系总表面积随盐浓度增加显著降低。盐浓度≤ 150 mmol· L-1时, 硅显著提高了高羊茅根系总表面积(P< 0.05); 当盐浓度≥ 200 mmol· L-1时, 硅对高羊茅根系总表面积无显著影响。盐浓度为0、50、100和150 mmol· L-1时, 添加硅较不添加硅高羊茅根系总表面积增幅分别为26.4%、29.5%、31.3%和29.7%。
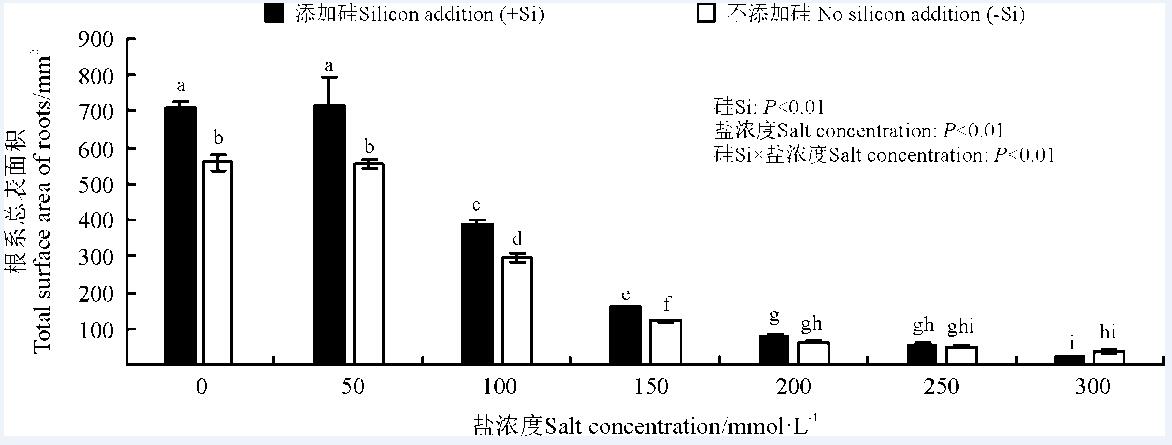 | 图3 不同盐浓度下硅对高羊茅根系总表面积的影响Fig. 3 Effect of the addition of silicon on the total surface of roots of tall fescue at different salt concentrations |
硅、盐浓度极显著影响了高羊茅根系体积(P< 0.01), 二者的互作对其无显著影响(P> 0.05)(图4)。高羊茅根系体积随着盐浓度增加逐渐降低(P< 0.05)。当盐浓度≤ 150 mmol· L-1时, 硅显著增加了高羊茅根系体积(P< 0.05)。
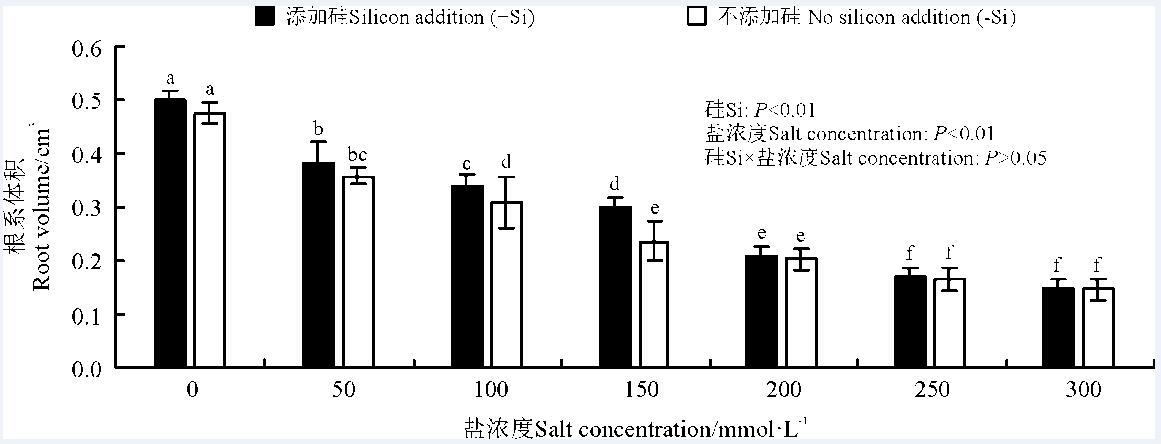 | 图4 不同盐浓度下硅对高羊茅根系体积的影响Fig. 4 Effect of the addition of silicon on the root volume of tall fescue at different salt concentrations |
硅、盐浓度及其交互作用对高羊茅根系平均直径无显著影响(P> 0.05)(图5)。随着盐浓度增加, 高羊茅根系平均直径有降低趋势(P> 0.05), 且添加硅对高羊茅根系平均直径有增加趋势(P> 0.05)。
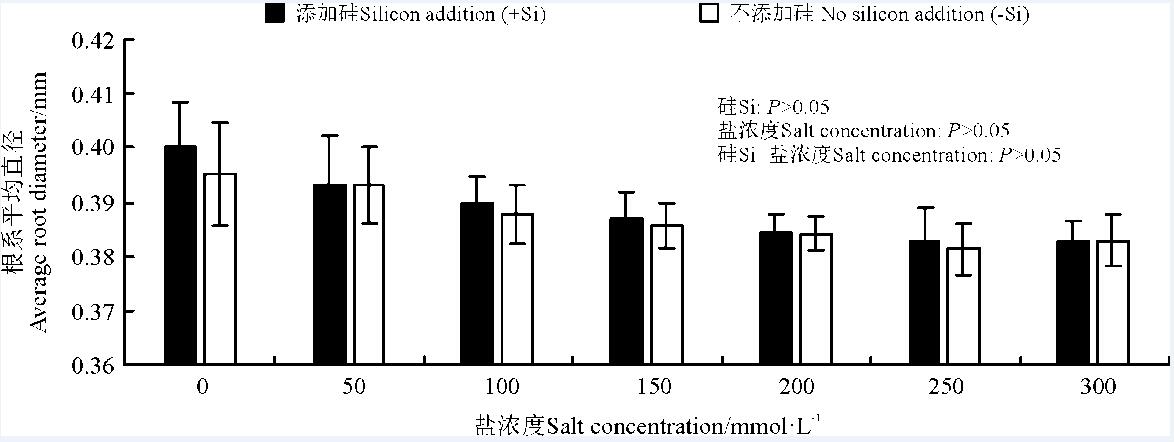 | 图5 不同盐浓度下硅对高羊茅根系平均直径的影响Fig. 5 Effect of the addition of silicon on the average root diameter of tall fescue at different salt concentrations |
硅、盐浓度均极影响了高羊茅根系总长(P< 0.01), 二者的互作对其有显著影响(P< 0.05)(图6)。随着盐浓度增加, 高羊茅根系总长逐渐减少, 盐浓度≤ 250 mmol· L-1时, 高羊茅根系总长随盐浓度增加显著降低(P< 0.05)。盐浓度≤ 150 mmol· L-1时, 硅显著提高了高羊茅根系总长(P< 0.05); 当盐浓度≥ 200 mmol· L-1时, 硅对高羊茅根系总长无显著影响; 盐浓度为150 mmol· L-1时, 添加硅较不添加硅高羊茅根系总长增幅最大, 为36%。
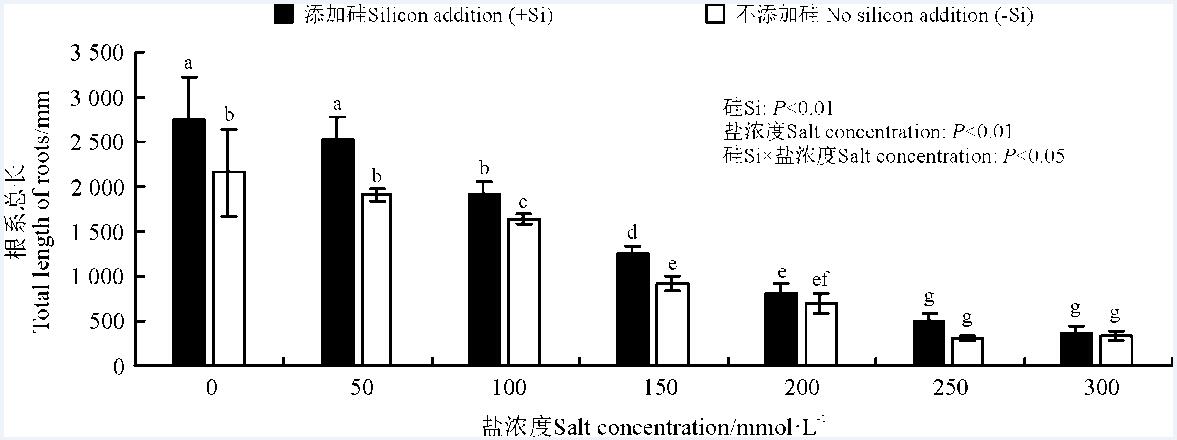 | 图6 不同盐浓度下硅对高羊茅根系总长的影响Fig. 6 Effect of the addition of silicon on the total length of roots of tall fescue at different salt concentrations |
在盐生境植物中受危害时, 其最直接的受害部位是根系, 而生物量作为植物耐盐性的直接指标之一, 是植物对盐害反映的综合体现[4, 20]。本研究表明, 虽然随盐浓度增加, 高羊茅根系生物量呈显著降低趋势, 但添加外源性矿质元素硅, 提高了高羊茅在盐浓度≤ 150 mmol· L-1时的根系生物量, 减轻了盐渍化对高羊茅根系的伤害效应, 促进了高羊茅根系在盐生环境中的生长, 增强了高羊茅适应盐生境的能力, 这与小麦[15]、玉米[17]在盐胁迫下对硅的响应相一致。说明添加外源性硅能够提高高羊茅根系吸水能力, 维持水分代谢平衡, 降低Na+在根系的积累, 减少盐分向地上部分的运输, 提高根系细胞活性, 促进了高羊茅在盐生环境下的生长。
根冠比的增加, 提高了植物对水分和养分的吸收及利用, 增强了植物的抗逆性, 促进了植物对外界不良环境的适应[4, 21]。本研究表明, 高羊茅根冠比随盐浓度增大呈现先上升后下降的趋势, 说明在较低盐浓度的生境下, 高羊茅根系较地上部分对盐的感应程度降低了, 这样有助于根系在盐生环境中吸收利用水分和养分[8], 维持高羊茅健康生长; 而在较高的盐生环境下, 高羊茅根系受到严重伤害, 生长受到抑制。硅显著增加了高羊茅的根冠比, 改变了地上部分和根系的分配模式, 这与黄瓜根冠比响应盐胁迫和外源性硅的过程类同[16], 说明盐生境下硅降低了高羊茅根系较地上部分对盐感应的程度, 提高了高羊茅根系在盐生境中吸收水分和矿物质的能力, 降低盐渍化环境对高羊茅的盐害程度, 促进其在盐渍环境下的生长, 提高了高羊茅的耐盐性。
根系是决定植物根量大小、适应盐生环境、吸收和利用土壤养分最直观的适应特征之一[4]。本研究中, 高羊茅根系表面积、体积、平均直径和总长均随盐浓度增加而呈降低趋势, 说明不同盐浓度生境对高羊茅根系形态特征有抑制作用[4]。在盐生环境下添加硅对根系平均直径有增加趋势; 当盐浓度≥ 200 mmol· L-1时, 硅对高羊茅根系表面积、体积和总长无显著影响, 但当盐浓度≤ 150 mmol· L-1时, 添加外源性硅显著提高了高羊茅根系表面积、体积、总长度; 在同一盐浓度下, 添加硅均在不同程度上提高了根系表面积、体积、总长, 说明盐生境下外源硅促进高羊茅根系的生长受盐浓度的限制, 在盐浓度较低时, 添加硅降低了高羊茅的盐害程度, 扩大了根系的分布, 促进了根系生长; 而盐浓度较高时, 添加硅对高羊茅根系影响不显著; 在盐浓度≤ 150 mmol· L-1时, 添加外源性硅通过增强高羊茅根系细胞活性, 提高细胞壁的扩展性, 增强根系活力, 增加高羊茅根系总长、体积和表面积, 促进高羊茅须根数量的增加, 增强高羊茅吸收水分和矿质元素的能力, 提高了高羊茅的耐盐性, 这与添加硅对马齿苋[8]、玉米[17]、番茄[18]的影响结果一致。因此, 添加矿质元素硅有利于高羊茅在一定盐生境下对水分、养分和微量元素的吸收, 增加高羊茅根系生物量, 促进根系生长, 增强高羊茅在盐生环境的生长能力。
(责任编辑 武艳培)
The authors have declared that no competing interests exist.
| [1] |
|
| [2] |
|
| [3] |
|
| [4] |
|
| [5] |
|
| [6] |
|
| [7] |
|
| [8] |
|
| [9] |
|
| [10] |
|
| [11] |
|
| [12] |
|
| [13] |
|
| [14] |
|
| [15] |
|
| [16] |
|
| [17] |
|
| [18] |
|
| [19] |
|
| [20] |
|
| [21] |
|