




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
野生赖草和垂穗披碱草抗旱能力比较
[张卫红1 , 刘大林1  , 苗彦军
, 苗彦军2 , 徐雅梅2 , 陈鸣晖1 , 邵将1 ]
, 苗彦军|
|


第一作者:张卫红(1991-),男,甘肃陇西人,在读硕士生,研究方向为牧草资源开发与利用。E-mail:[email protected]
干旱是制约西藏牧草生长发育的一个重要因素,为此,本研究以西藏野生赖草( Leymus secalinus)和披碱草( Elymus nutans)作为试验材料,探究两种野生牧草抗旱能力的强弱,以期为西藏干旱半干旱区域生态恢复和栽培草地建植时草种的选择提供参考依据。结果表明,两种野生牧草的株高增长速率、植株含水量和叶绿素含量均随干旱时间的增加而降低( P<0.05);丙二醛和游离脯氨酸含量随着干旱胁迫时间的增加而增加( P<0.05)。赖草可溶性糖含量呈先减后增的变化趋势,而垂穗披碱草可溶性糖含量则呈减-增-减的变化趋势;可溶性蛋白含量均表现出先减后增的变化趋势。利用隶属函数{正相关函数模型 F1 =( Xi-Xmin)/( Xmax- Xmin)和负相关函数模型 F2 =[1 -( Xi-Xmin)] /( Xmax- Xmin)}法对两种野生牧草的两个生物学特性指标(植株含水量和株高)和5个生理特性指标(叶绿素、游离脯氨酸、丙二醛、可溶性糖和可溶性蛋白)进行综合评价,得出赖草的抗旱能力大于垂穗披碱草。
Drought is one of the most important factors restricting the growth and development of grasslands in Tibet. To counter these effects, we studied drought resistance ability in two species of wild forage plant, by collecting seeds of Leymus secalinus and Elymus nutans in Tibet, in order to provide reference for the ecological restoration of arid and semi-arid areas, and selection of grass species in artificial grassland. The results showed that the growth rate, plant water content, and chlorophyll content of two wild forage species decreased with the increase of drought time, and the difference was significant ( P<0.05). The contents of malondialdehyde and free proline increased with increase in drought time, and the difference was significant ( P<0.05). Soluble sugar content of L. massoniana decreased first and then increased, while the soluble sugar content of E. nutans decreased first and then increased, and finally decreased. The soluble protein content decreased first and then increased. The two biological characteristics and the five physiological characteristic indexes of the two wild forage species were comprehensively evaluated by the membership function method, and the drought resistance of L. secalinus was greater than that of E. nutans.
西藏作为我国五大牧区之首, 畜牧业一直是当地农业发展的支柱产业, 是当地农牧民的主要经济来源[1]。长期以来, 西藏天然草地不仅为当地农牧民提供生产、生活的物质基础, 还是我国重要的生态屏障和主要水资源发源地[2, 3]。牧草作为畜牧业实现健康可持续发展的基础, 其产量的高低直接决定着畜牧业发展水平[4]。但由于西藏寒冷、干旱条件的限制[5, 6], 加之过度放牧、滥伐滥挖和鼠害等综合因素的影响, 草地产草量低, 退化日益严重[7]。科技工作者关于西藏草地生态和牧草抗寒性研究等方面做了大量工作[8, 9, 10], 但关于西藏当地牧草抗旱性的研究却并不多见。干旱主要发生在西藏北部的广大地区, 在该区域每年每个季节干旱都有发生; 但藏南地区为季风气候, 干湿季明显, 每年10月至次年3月干燥少雨。由此可见, 干旱是制约西藏天然草地牧草生长发育的一个重要因素[11], 干旱往往会影响植物的蒸腾、水势、吸收和新陈代谢, 以及酶促反应、休眠和生长发育, 从而降低牧草生产力, 使草畜矛盾尖锐, 严重限制当地畜牧业的可持续发展[12]。
赖草(Leymus secalinus)和垂穗披碱草(Elymus nutans)作为西藏天然草地重要的乡土植物, 具有营养价值高、适口性好和适应性强等诸多优良特性, 在西藏畜牧业生产和生态恢复治理中具有十分广阔的应用前景[13, 14, 15]。相关学者以PEG-6000作干旱渗透介质对披碱草种子的萌发和幼苗生长特性进行了研究[16]。采用盆栽法对垂穗披碱草在干旱胁迫下的各项生理指标进行了测定[17], 对干旱胁迫下6种披碱草属野生牧草苗期生理变化进行了研究[18], 但目前关于赖草抗旱性方面的报道较少, 尤其关于这两个牧草抗旱性能力强弱比较的研究还未见详细报道。苗期是研究植物对逆境胁迫最敏感时期[19], 鉴于此, 本研究以采集于西藏当地的野生赖草和垂穗披碱草种子作为试验材料, 对干旱胁迫下两种牧草苗期生物学形态特性和生理指标进行比较, 旨在为西藏干旱、半干旱区生态恢复和栽培草地建植提供科学理论依据。
试验材料为2013年采集的赖草和垂穗披碱草种子。赖草种子采集于西藏达孜县有卵石分布的阳坡草地, 当地优势物种是高山嵩草(Kobresia pygmaea), 海拔高度为4 189 m, 年降水量约444 mm, 年无霜期约130 d。垂穗披碱草种子采集于申扎县高寒草甸, 当地植被以青藏苔草(Carex moorcroftii )和矮嵩草(K. humilis)为主, 海拔高度为4 852 m, 年降水量约298.6 mm, 年均温0.4 ℃。通过人工采集牧草种子, 用剪刀将牧草成熟的穗剪下, 装入布袋, 带回实验室, 在日光下自然风干, 并进行清选, 将清选干净后的种子放入4 ℃低温保存。
试验于西藏农牧学院高寒草地实验室进行。采用盆栽法, 花盆内直径为30 cm, 高25 cm。土壤采集于西藏农牧学院草业科学教学实习基地, 砂壤土, pH 5.77, 有机质10.87 g· kg-1, 全氮1.05 g· kg-1, 碱解氮0.16 g· kg-1, 速效钾41.60 mg· kg-1, 速效磷19.67 mg· kg-1。去除土壤中的石子、枯枝落叶和植物根茎, 并将其置于105 ℃烘箱中烘5 h, 待自然冷却后在每千克土壤中添加尿素10 g, 混合均匀后在每个花盆中加入10 kg土壤, 然后在每个花盆内分别播种赖草种子400粒和垂穗披碱草种子300粒, 行距为1 cm, 播深1.5 cm, 浇水至各个花盆重量相等, 每种草5次重复, 共计10盆。将花盆放入人工智能气候培养箱中进行培养, 培养条件为12 h的15℃黑暗处理和12 h的25 ℃光照处理、光强为4 000 Lx。对所有花盆正常浇水, 待苗长出2片叶子时进行间苗, 保证每盆200株幼苗, 待牧草长到3片叶片时, 作为对照进行一次采样, 样品迅速用锡纸包裹, 液氮迅速冷冻后, 放入-80 ℃低温冰箱保存[20], 测其各项指标, 每个指标3次重复。然后不再进行浇水, 每隔5 d采样一次, 干旱处理周期为20 d。
土壤含水量和植株含水量采用烘干法。土壤含水量是将采集的土壤105 ℃烘干至恒重, 减少的土壤质量与烘干前质量的比值; 植株含水量是105 ℃杀青30 min后, 65 ℃烘干至恒重, 减少的质量与鲜样质量的比值。用直尺测量株高(绝对高度), 精确到0.1 cm, 每盆测5株; 游离脯氨酸采用茚三酮法[21]; 叶绿素采用95%酒精浸泡法测定[22]; 丙二醛采用硫代巴比妥酸法[23]; 可溶性糖采用苯酚法[23]; 可溶性蛋白采用考马斯亮蓝G-250法[23]。
抗旱性评价采用隶属函数法[24], 以两种野生牧草在不同干旱胁迫下的株高、含水量、游离脯氨酸、叶绿素、丙二醛、可溶性糖和可溶性蛋白作为评价指标进行综合评价。计算公式:
F1=(Xi-Xmin)/(Xmax-Xmin).
式中:Xi表示第i个指标值, Xmax表示所有品种第i个指标的最大值, Xmin表示所有品种第i个指标的最小值。
若所测指标与植物的耐旱性呈负相关, 则用反隶属函数计算其抗旱隶属函数值。计算公式:
F2= [1-(Xi-Xmin)]/(Xmax-Xmin).
根据两种野生牧草各指标的具体隶属函数值, 计算出平均值后进行比较, 平均值越大表示该牧草的抗旱性越强[25]。
采用Excel 2013进行数据输入和预处理。然后用SPSS 19.0中的单因素方差分析(One-way ANOVA)对同种牧草在不同胁迫天数下的各指标对进行方差分析, Duncan法对其多重比较; 独立样本T检验对相同胁迫天数下土壤含水量和不同牧草间的各指标进行差异性分析。最后用Excel 2013作图。
随着干旱胁迫时间的增加, 土壤含水量逐渐降低, 赖草土壤含水量下降幅度小于垂穗披碱草(表1); 干旱胁迫5 d, 两种野生牧草的土壤含水量差异不显著(P> 0.05), 而干旱胁迫时间≥ 10 d时, 赖草土壤含水量显著高于垂穗披碱草(P< 0.05)。干旱胁迫初期土壤水分散失由土壤和植物蒸腾共同作用造成, 而干旱胁迫后期, 主要由植物蒸腾散失造成土壤水分的散失[26]。由此可见, 两种牧草在土壤特性一致的情况下, 土壤水分变化是由不同植物蒸腾散失引起的, 其中, 赖草对土壤水分蒸发量的影响小于垂穗披碱草, 具有较好的保水能力。
| 表1 干旱胁迫各时期土壤含水量(%)变化 Table 1 Changes of soil water content(%) of drought stress in different drought stress period |
随着干旱胁迫天数的增加, 两种牧草植株的含水量均降低(图1)。植株绝对株高却随着干旱胁迫时间的增加而增加(P< 0.05), 但株高增加量随着干旱时间的增加而降低。干旱胁迫条件下野生赖草和野生垂穗披碱草植株含水量和株高均差异显著(P< 0.05)。干旱胁迫至20 d时, 赖草含水量较CK降低了29.52%, 而垂穗披碱草含水量却降低了37.42%; 赖草株高增加了23.82 cm, 垂穗披碱草株高仅增加了18.22 cm。由此可见, 干旱胁迫对垂穗披碱草植株形态特征的影响大于赖草。
 | 图1 干旱胁迫下苗期植株形态特征比较 注:不同大写字母表示相同胁迫天数不同牧草间差异显著(P< 0.05), 不同小写字母表示同种牧草在不同胁迫天数下差异显著(P< 0.05)。下同。Fig. 1 Comparison of the morphological characteristics of seedlings under drought stress Note: Different capital letters indicate signification between forages under the same drought stress days at the 0.05 level, and different lowercase letters indicate signification among different drought stress days in the same forage at the 0.05 level. similarly for the following figures. |
2.3.1 叶绿素含量比较 叶绿素是植物叶片进行光合作用的重要色素, 其含量越高, 光能利用效率越高[27, 28]。两种牧草叶片中的叶绿素含量随着干旱胁迫时间的增加呈现递减趋势, 且降幅也随干旱胁迫时间的增加而降低(图2)。干旱胁迫20 d, 赖草叶片中叶绿素较CK降低了12.64 mg· g-1(P< 0.05), 而垂穗披碱草叶片中叶绿素含量较CK下降了9.99 mg· g-1(P< 0.05), 从叶绿素减少量分析, 干旱胁迫对赖草叶绿素含量影响较大, 对赖草叶片叶绿素合成的抑制效果大于对垂穗披碱草的。但干旱胁迫整个过程中, 赖草叶片中叶绿素含量均大于垂穗披碱草(P< 0.05)。
 | 图2 干旱胁迫下叶绿素含量比较Fig. 2 Comparison of chlorophyll content under drought stress |
2.3.2 游离脯氨酸含量比较 牧草体内游离脯氨酸的积累是植物受到逆境胁迫时, 表现较为敏感的一种渗透调节物质[29], 具有保护牧草生物大分子和维持膜结构等重要作用[23], 其含量的高低可以反映牧草植物对干旱胁迫的抗性能力的强弱[26, 30, 31]。两种牧草的游离脯氨酸含量随着干旱胁迫时间的增加而呈递增趋势(图3), 干旱胁迫0 d(CK)时, 赖草和垂穗披碱草叶片中游离脯氨酸含量差异不显著(P> 0.05), 干旱胁迫5 d时, 游离脯氨酸增速较缓, 干旱胁迫5 d以后, 则迅速增加, 且赖草体内游离脯氨酸含量显著高于垂穗披碱草(P< 0.05)。干旱胁迫至20 d时, 赖草体内游离脯氨酸含量较CK增加了2 219.1 μ g· g-1(P< 0.05), 而垂穗披碱草较CK仅增加了1 338.53 μ g· g-1(P< 0.05), 是垂穗披碱草的1.66倍(P< 0.05)。由此可见, 赖草体内游离脯氨酸对干旱胁迫的应激反应大于垂穗披碱草。
 | 图3 干旱胁迫下游离脯氨酸含量比较Fig. 3 Comparison of proline content under drought stresss |
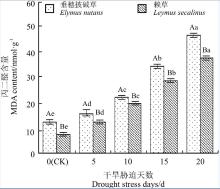
2.3.3 丙二醛含量比较 丙二醛(MDA)是植物在逆境胁迫下发生膜脂过氧化作用的终产物之一, 其在植物体内的积累会对膜和细胞进一步造成伤害, MDA含量越高, 表明植物组织受到的伤害程度越重[32, 33]。两种牧草中MDA的含量随着干旱胁迫时间的增加而增加(图4), 不同牧草在不同干旱胁迫阶段的响应有所不同。垂穗披碱草中MDA含量的增加幅度大于赖草。干旱胁迫20 d时, 垂穗披碱草的MDA含量较CK增加了35.01 nmol· g-1(P< 0.05), 而赖草增加了30.43 nmol· g-1(P< 0.05), 在整个干旱胁迫处理过程中, 垂穗披碱草中MDA含量均显著高于赖草(P< 0.05)。因此, 无论从干旱胁迫过程中MDA的增加量, 还是MDA总含量进行分析, 干旱胁迫对垂穗披碱草的损害程度大于赖草。
 | 图4 干旱胁迫下丙二醛含量比较Fig. 4 Comparison of MDA content under drought stress |
2.3.4 可溶性糖含量比较 可溶性糖(SS)是植物在干旱胁迫环境下体内积累的一种渗透调节物质, 可提高植株幼苗的吸水能力及降低叶片的渗透势, 也是合成其它有机溶质的碳架和能量来源[33]。随着干旱胁迫时间的增加, 垂穗披碱草SS含量呈减-增-减的变化趋势。赖草呈先减小后增大的趋势, 不同牧草SS含量对干旱胁迫的响应也有所不同, 干旱胁迫初期(5和10 d), 赖草SS含量高于垂穗披碱草(P< 0.05), 随着干旱胁迫时间的延长(15和20 d), 垂穗披碱草体内SS含量显著高于赖草(P< 0.05)。
 | 图5 干旱胁迫下可溶性糖含量比较Fig. 5 Comparison of soluble sugar content under drought stress |
2.3.5 可溶性蛋白含量比较 植物体内的可溶性蛋白(SP)大多数是参与植物各种代谢的酶类, 测其含量是了解植物体总代谢的一个重要生理生化指标[34]。垂穗披碱草随着干旱胁迫时间的增加呈增-减-增的变化趋势(图6); 而赖草随着干旱胁迫时间的增加呈先减小后增大的变化趋势, 其中赖草在干旱胁迫5 d时较对照下降了29.74 μ g· g-1(P< 0.05), 达到干旱处理最低值。干旱胁迫5、10和15 d时, 垂穗披碱草SP含量均大于赖草(P< 0.05); 而CK和干旱胁迫20 d时, 赖草SP含量大于垂穗披碱草(P< 0.05); 干旱胁迫20 d时, 垂穗披碱草较CK的SP含量增加了48.85 μ g· g-1(P< 0.05), 而赖草却增加了49.74 μ g· g-1(P< 0.05)。由此可见, 干旱胁迫对赖草和垂穗披碱草SP含量的影响不尽相同。
 | 图6 干旱胁迫下可溶性蛋白含量比较Fig. 6 Comparison of soluble protein content under drought stress |
采用隶属函数法对干旱胁迫下上述两种野生牧草苗期的两个生物学特性指标(植株含水量和株高)及5个生理特性指标(叶绿素、游离脯氨酸、丙二醛、可溶性糖和可溶性蛋白)进行综合评价。结果表明, 赖草综合评价值较大, 为0.55, 而垂穗披碱草的为0.45, 表明, 赖草的抗旱能力大于垂穗披碱草(表2)。
| 表2 野生赖草和野生垂穗披碱草抗旱能力综合比较 Table 2 Comprehensive comparison of drought resistance of Elymus nutans and Leymus secalinus |
土壤水分是植物生长的一个重要因素, 对于植物的生理活动起着至关重要的作用[35, 36]。本研究结果表明, 干旱胁迫过程中, 两种草的土壤含水量下降程度不同, 在外界环境一致的情况下, 其原因为不同植物的蒸腾和水分散失能力的差异性所造成土壤水分差异。其中种植赖草的土壤含水量下降程度小于垂穗披碱草, 表明赖草对土壤水分的散失量小于垂穗披碱草, 较垂穗披碱草具有较强的避旱能力。
赖草和垂穗披碱草牧草幼苗的株高增幅、植株含水量和叶绿素均随干旱胁迫时间的增加而呈现下降趋势, 游离脯氨酸和MDA含量随着干旱胁迫时间的增加而增加。其原因是牧草植物受到干旱胁迫后, 植物体光合作用受到抑制、水分利用率受到影响, 从而导致植物生长受到抑制[7]。同时, 植物在干旱胁迫环境下, 两种野生牧草幼苗受到了不同程度的伤害, 导致MDA含量的升高。植株为了抵抗干旱胁迫, 以提高游离脯氨酸含量来减轻干旱胁迫造成的伤害。该结果与干旱胁迫下对早熟禾(Poa annua)[24]、黑麦草(Lolium perenne)[37]、高羊茅(Festuca elata)[38]和垂穗披碱草[18]等禾本科牧草的研究结果一致。
可溶性糖和可溶性蛋白是植物重要的渗透调节物质[39]。本研究发现赖草可溶性糖含量呈先减小后增大的变化趋势, 而垂穗披碱草可溶性糖含量则呈减-增-减的变化趋势; 两种野生牧草中可溶性蛋白均呈先减小后增大的变化趋势。其原因可能是因为干旱初期植物主动适应逆境, 消耗一部分可溶性糖和可溶性蛋白进行植物体内的代谢调节或者可溶性糖和可溶性蛋白的合成受到了抑制[40], 故植物体内可溶性糖和可溶性蛋白含量减少, 待适应一段时间后, 体内便积累大量可溶性糖, 以减少逆境环境对植物的危害, 而到干旱胁迫5和10 d时, 植物适应干旱胁迫环境过程中当细胞内的可溶性糖和可溶性蛋白含量积累到一定量的时候会维持相对稳定[10]。但垂穗披碱草可溶性糖含量最后呈降低趋势, 其原因可能是干旱胁迫至15 d后, 植株体内水分亏损, 从而抑制可溶性糖的合成[41], 至于具体原因有待进一步研究。同时在研究中发现不同植物在不同的干旱胁迫条件下各项指标表现出不同的响应规律。其原因是两种牧草遗传背景不同, 故两种牧草对干旱胁迫的生理响应之间也就具有差异性, 这在不同种类牧草的抗旱性研究中已经得到证实[42, 43]。
综上所述, 随着干旱胁迫时间的增加, 不同牧草在同样干旱胁迫下的响应也不尽相同, 整体上赖草对干旱胁迫的抗性大于垂穗披碱草。本研究采用隶属函数法对西藏野生赖草和垂穗披碱草苗期在干旱胁迫下的两个生物学形态特征指标和5个生理指标进行综合分析对比, 得出赖草的抗旱能力大于垂穗披碱草。由此可见, 赖草在干旱和半干旱区生态恢复及栽培草地建植中具有重要用途。
(责任编辑 张瑾)
The authors have declared that no competing interests exist.
| [1] |
|
| [2] |
|
| [3] |
|
| [4] |
|
| [5] |
|
| [6] |
|
| [7] |
|
| [8] |
|
| [9] |
|
| [10] |
|
| [11] |
|
| [12] |
|
| [13] |
|
| [14] |
|
| [15] |
|
| [16] |
|
| [17] |
|
| [18] |
|
| [19] |
|
| [20] |
|
| [21] |
|
| [22] |
|
| [23] |
|
| [24] |
|
| [25] |
|
| [26] |
|
| [27] |
|
| [28] |
|
| [29] |
|
| [30] |
|
| [31] |
|
| [32] |
|
| [33] |
|
| [34] |
|
| [35] |
|
| [36] |
|
| [37] |
|
| [38] |
|
| [39] |
|
| [40] |
|
| [41] |
|
| [42] |
|
| [43] |
|