



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
基于无人机的青藏高原鼠兔潜在栖息地环境初步研究
[郭新磊1, 2 , 宜树华1  , 秦彧
, 秦彧1 , 陈建军1, 2 ]
, 秦彧|
|


第一作者:郭新磊(1990-),男,河北魏县人,在读硕士生,主要从事无人机生态学研究。E-mail:[email protected]
高原鼠兔( Ochotona curzoniae)是青藏高原生态系统中的关键种之一,其栖息地环境特征对其空间分布的影响一直是重要的研究内容。由于传统地面观测方法耗时耗力,而卫星遥感分辨率低没法识别,因而对高原鼠兔栖息地的研究多为定性且缺少大范围量化研究。无人机航拍为研究高原鼠兔栖息地提供了新的可能,本研究于2015年6月-8月在青藏高原进行了定点定高航拍,共布设了约300个工作地点,获取了约1 800张航拍照片;每张照片覆盖地面26 m×35 m,每个像元覆盖地面约为1 cm2;通过自主开发的照片处理软件对鼠洞进行自动识别和人工校正,并结合样地的地表温度、植被、土壤水分等信息进行分析。结果表明,1)高原鼠兔洞口密度在不同草地类型中的分布显著不同( P<0.05),高寒草甸高原鼠兔洞口密度显著高于其它草地类型,荒漠高原鼠兔洞口密度最小;2)高原鼠兔最适宜的栖息地环境:NDVI值为0.5~0.6,土壤含水量为20%~25%,生长季陆地表面温度为28.5~29 ℃;3)高原鼠兔洞口密度与NDVI和土壤水分呈显著正相关关系( P<0.05),但与陆地表面温度呈显著负相关关系( P<0.05)。以上结果说明,植被和土壤水热是影响高原鼠兔选择栖息地的主要因子,高原鼠兔偏向于选择植被条件较好、土壤含水量较高,生长季陆地表面温度较低的区域。
Plateau pikas ( Ochotona curzoniae) were considered a keystone species on the Qinghai-Tibetan plateau (QTP). Much attention has been paid to the effect of habitat environment on their spatial distribution. However, most of the previous studies mainly focused on qualitative description, because pikas’ holes were difficult to identify by satellite-based remote sensing and traditional field observation was time-consuming. High-precision photography and low costs make the use of unmanned aerial vehicles (UAVs) a feasible option for investigating the spatial distribution of pikas at the plot or even the regional scale. In the present study, we investigated pikas’ burrow density by using aerial photography with light UAVs from July to August 2015. Altogether, approximately 300 work points were set and 1800 aerial photos were obtained. Each photograph covered 26 m×35 m on the ground and each pixel covered approximately 1 cm2. Coupled with normalized difference vegetation index (NDVI), soil water content (SWC) and land surface temperature (LST) data derived from satellite——based remote sensing, we analysed the effect of habitat environment on plateau pikas’ spatial distribution. The results showed that 1) the burrow density of pikas significantly varied with grassland types ( P<0.05); alpine meadows had the maximum burrow density, whereas deserts had the lowest; 2) the most suitable habitat for pikas was 0.5~0.6 for NDVI, 20%~25% for SWC, and 28.5~29 ℃ for LST, respectively; and 3) burrow density was positively correlated with NDVI and SWC ( P<0.05), but significantly negatively correlated with LST ( P<0.05). Our results suggested that NDVI, SWC, and LST were the dominant factors affecting the distribution of pikas. Owing to the high resolution of the aerial photographs, UAVs provided a novel and efficient method of improving our understanding of the role of pikas over large regions on the QTP.
草地是地球上广泛分布的生态系统类型之一, 大约占据了陆地表面积的20%, 在全球碳循环中起着重要作用[1]。中国草地主要分布在青藏高原、内蒙古高原、东北平原、黄土高原以及新疆等地。其中, 青藏高原草地面积大约为1.63× 106 km2, 高寒草地是其重要的植被类型, 约占总面积的2/3, 是中国乃至世界的重要牧区之一。然而, 大约1/3的高原草地在近几十年内发生了不同程度的退化[2]。除了过度放牧[3]、气候变化和多年冻土退化[4, 5], 高原鼠兔(Ochotona curzoniae)被认为是高寒草地退化的重要因素之一。
高原鼠兔为兔形目鼠兔科鼠兔属动物, 广泛分布于我国青海、西藏、甘肃、四川西北部, 以及尼泊尔、巴基斯坦东部等地[6]。由于其所挖掘的洞穴可为其它生物诸如鸟类等提供住所[7], 改变土壤的微地形提高物种丰富度, 增加水分的入渗率并减少地表径流[8], 自身几乎是所有高原上捕食者的重要食物来源, 同时对生态系统物质循环起着积极作用[9], 因此被认为是青藏高原生态系统中的关键种[10]。但是, 高原鼠兔种群密度的爆发和草地退化也有着较高的一致性, 因而有研究认为高原鼠兔的存在是青藏高原地区草地退化的主要原因之一[11, 12], 并在一些地区开展了大规模的灭鼠行动[13]。鉴于目前对高原鼠兔在高寒草地生态系统中的作用认识还存在着很大的分歧, 需要大力开展相关研究, 进而制定出合理的草地退化减缓措施。高原鼠兔的栖息地环境特征的研究一直方兴未艾[14, 15, 16], 研究其空间分布特征和栖息地环境有助于提高对高原鼠兔在生态系统中的作用[17]以及环境如何影响高原鼠兔的分布等方面的认识, 然而受限于高效获取鼠兔种群密度的方法, 目前已开展的绝大多数研究只是局限于样地尺度, 缺乏大尺度如高原尺度关于高原鼠兔和栖息地环境要素的空间分布格局的报道, 而且以定性研究为主, 缺少定量分析的研究。
传统的获取高原鼠兔种群数量的方法有标志重捕法[14]、步行计数法(walk-transects)[18]和洞口计数法, 即获取地面总洞口, 结合堵洞盗洞法[7]获得有效洞口数, 再结合少量捕捉法, 获得相应的转换系数。此类方法的优势在于计数和对微生境描述较精确[16], 但是由于其所需较多的人力、物力, 无法推广到大尺度的高原鼠兔和栖息地环境要素的分布格局的研究中。卫星遥感虽然可以提供大尺度的环境要素的信息, 但是由于分辨率太低, 无法提供高原鼠兔的洞口分布数据。高原鼠兔主要栖息于植被高度较低的平坦开阔的生境中[14, 19], 因而无人机航拍可能成为高原鼠兔种群密度研究的新手段。近几年, 由于智能手机中的传感器(比如GPS、陀螺仪)的小型化和普及, 无人机的成本急剧下降, 使得大范围应用无人机进行草地生态研究成为可能[20]。鉴于此, 本研究通过无人机航拍调查高原鼠兔洞口密度在青藏高原不同草地类型的分布特征, 并结合植被、土壤水分、地表温度等资料初步分析高原鼠兔的栖息地特征, 以期为青藏高原地区生态系统的科学管理和保护提供理论依据。
青藏高原位于中国西南部, 横跨31个经度(73° 18'52″-104° 46'59″ E)、纵贯13个纬度(26° 00'12″-39° 46'50″ N), 东西长2 945 km, 南北宽1 532 km, 面积为2.57× 106 km2。青藏高原地形复杂, 海拔变化大(100~8 846 m), 平均海拔超过4 000 m, 地面气温区域差异显著, 温度季节差异明显。行政区划上, 青藏高原涉及西藏自治区、青海省绝大部分地区, 以及云南省、四川省、甘肃省和新疆部分地区[21, 22]。本研究的区域主要为西藏自治区与青海省。研究区域内植被类型主要包含高寒草甸(alpine meadow)、灌丛(shrub)、沼泽(swamp)、草原化草甸(steppe meadow)、退化草甸(degraded meadow)、退化沼泽(degraded swamp)、草原(steppe)和荒漠(desert)8种类型, 分别占总采样面积的23.85%、1.83%、18.35%、6.42%、8.72%、2.29%、36.70%和1.83%。
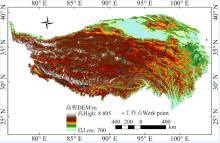
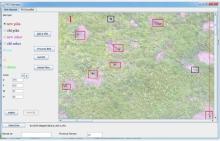
1.2.1 高原鼠兔洞口密度数据获取 2015年7月24日-8月24日, 在青藏高原开展了航拍工作, 共布设了约300个工作地点(图1), 每个工作点设置1~2个200 m× 100 m航线, 每个航线每隔50 m设置一个航点, 无人机自主飞行到航点进行拍摄。航线超过500个, 共获取了约1 800张航拍照片。本研究使用的无人机的拍摄高度为20 m, 每张照片覆盖的地表面积约为26 m× 35 m, 航拍照片的像素为1 200万, 每个像素的分辨率大约为1 cm, 而高原鼠兔的洞口的直径大约为5 cm, 在航空照片上可以清晰辨识洞口(图2)。然后通过自主开发的软件手工画分洞口(图2), 获取每张照片上的鼠洞密度[23]。
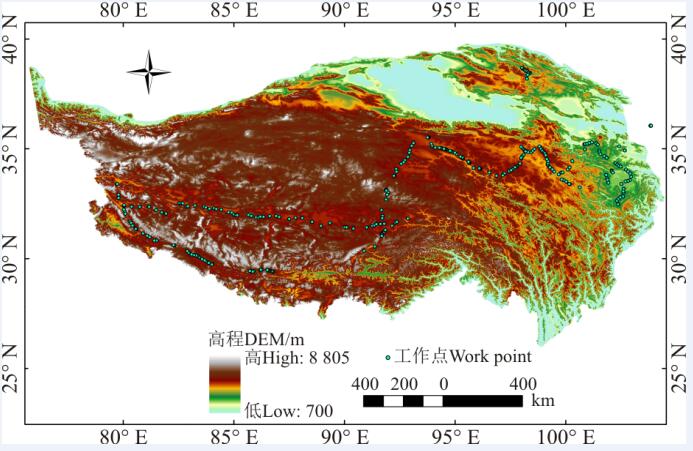 | 图1 青藏高原野外航拍工作点Fig. 1 Work points set on the Qinghai-Tibetan Plateau for aerial photographing in field |
 | 图2 手工勾画航空照片中的高原鼠兔洞口Fig. 2 Outline of the burrows of plateau pikas in the aerial photographs |
1.2.2 环境数据来源
1)NDVI数据来源:本研究NDVI使用的是MOD13Q1产品数据, 其时间分辨率为16 d、空间分辨率为250 m。该植被产品经过了大气校正, 并对水域、云、气溶胶和云阴影的影响进行了掩膜处理。MOD13Q1产品数据从USGS(United States Geological Survey) 网站(http://glovis.usgs.gov/)上下载。综合使用ArcGIS和ENVI软件以及MODIS数据预处理工具(MRT等)完成青藏高原区域NDVI数据的拼接、裁剪等处理, 获得2001-2012年生长季(6月-8月)NDVI平均值[23]。然后, 提取各航点的NDVI值, 用来分析鼠兔洞口分布与NDVI之间的关系。
2)陆地表面温度(land surface temperature)数据来源:陆地表面温度数据使用的是MOD11A2数据产品, 是每8日的地温最大值合成数据, 原始数据为MOD11A1(逐日地温产品)数据。综合使用ArcGIS和ENVI软件以及MODIS数据预处理工具(MRT等)完成青藏高原区域MODIS LST数据的预处理, 包括图像拼接与裁剪, 投影转换(WGS84), 波段运算(比例因子为0.02), 16 d最大值合成等, 获得2001-2012年生长季(6月-8月)LST平均值[24]。然后, 提取各航点的LST值, 用来分析高原鼠兔洞口密度与陆地表面温度(LST)之间的关系。
3)土壤水分(soil water content)数据来源:土壤水分数据来自于中国科学院青藏高原研究所阳坤研究员团队[25, 26], 空间分辨率为0.25° × 0.25° , 时间分辨率为逐日, 时间序列为2002-2010年[23]。计算2002-2010年多年生长季(6月-8月)平均值。然后, 提取各航点的土壤水分数据, 用来分析高原鼠兔洞口密度与土壤含水量(SWC)之间的关系。
采用SPSS 17.0软件对不同草地类型、不同NDVI区间、不同地表温度区间和不同土壤水分区间高原鼠兔洞口密度进行单因素方差分析(ANOVA), 再采用最小显著极差法(Least Significant Difference)在0.05水平上检验其差异显著性; 鼠兔洞口密度与NDVI, 地表温度和土壤含水量的相互关系采用线性回归分析; 所有图表采用Origin 9.0软件进行绘制。
总体来看, 高原鼠兔洞口密度大小依次为高寒草甸> 草原化草甸> 退化草甸> 退化沼泽> 沼泽> 草原> 灌丛> 荒漠(图3)。其中, 高寒草甸中高原鼠兔洞口密度最高, 为481.61个· hm-2, 草原化草甸为241.71个· hm-2, 退化草甸为206.73个· hm-2, 退化沼泽为191.24个· hm-2, 沼泽为111.69个· hm-2, 草原为69.05个· hm-2, 灌丛为17.63个· hm-2, 荒漠最低, 为7.71个· hm-2。
 | 图3 高原鼠兔洞口密度在不同草地类型中的分布特征 注:AM, 高寒草甸; SH, 灌丛; SW, 沼泽; SM, 草原化草甸; DM, 退化草甸; DS, 退化沼泽; ST, 草原; DE, 荒漠; 。Fig. 3 Density distribution of plateau pika burrows in different grassland types Note: AM, alpine meadow; SH, shrub; SW, swamp; SM, steppe meadow; DM, degraded meadow; DS, degraded swamp; ST, steppe; DE, desert. |
研究区域内所有像元的NDVI值都介于0.1~0.6, 以0.1为间隔, 将NDVI值依次划分为0.1~0.2、0.2~0.3、0.3~0.4、0.4~0.5和0.5~0.6共5个区间。各NDVI区间内的高原鼠兔洞口的密度依次为268.02、206.70、353.41、17.58和481.65个· hm-2(图4)。回归分析进一步指出, 高原鼠兔洞口密度与NDVI值显著正相关(y=735.55x-55.77, R2=0.41, P< 0.05)(图4), 表明高原鼠兔的洞口密度随着NDVI值的增加而增大。
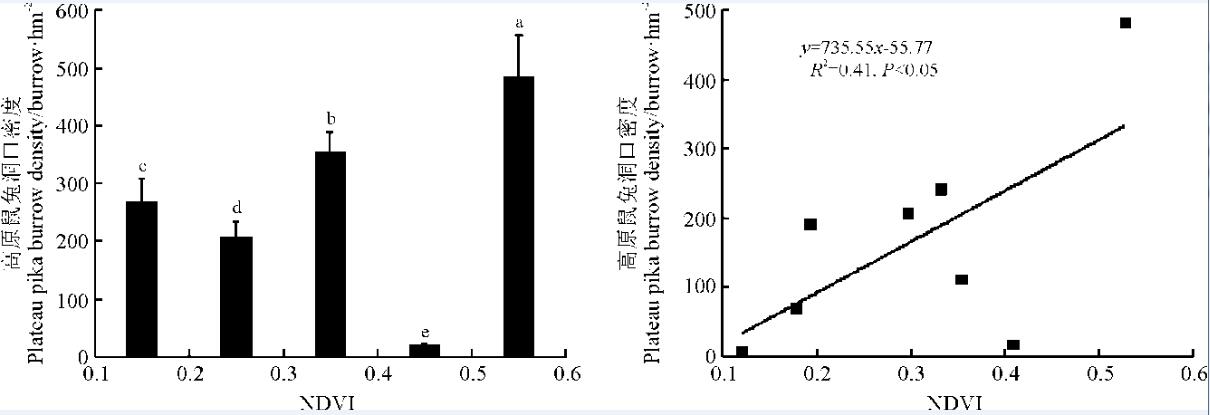 | 图4 高原鼠兔洞口密度在不同NDVI区间的分布特征及其与NDVI的关系Fig. 4 Plateau pika burrow densities in different NDVI interval and its relationship with NDVI |
研究区域内, 土壤含水量的分布范围为5%~25%, 以5%为间隔, 将土壤含水量依次划分为5%~10%、10%~15%、15%~20%和20%~25%共4个区间。各个水分区间内的高原鼠兔洞口的密度依次为7.69、86.7、432.86和800.11个· hm-2(图5)。回归分析结果显示, 高原鼠兔洞口密度与土壤含水量显著正相关(y=2 566.9x-2 364.31, R2=0.52, P< 0.05)(图5), 即高原鼠兔的洞口密度随着土壤含水量的增加而增大。
 | 图5 高原鼠兔洞口密度在不同土壤水分区间的分布特征及其与土壤含水量的关系Fig. 5 Plateau pika burrow densities in different soil water content (SWC) interval and its relationship with SWC |
研究区域内陆地表面温度介于28.5~30.5 ℃, 以0.5 ℃为间隔, 将陆地表面温度值依次划分为28.5~29.0 ℃、29.0~29.5 ℃、29.5~30.0 ℃和30.0~30.5 ℃共4个区间。在陆地表面温度为28.5~29.0 ℃的区间内, 高原鼠兔的洞口密度为1 232.97个· hm-2; 在地表温度为29.0~29.5 ℃的区间内, 高原鼠兔的洞口密度为17.58个· hm-2; 在陆地表面温度为29.5~30.0 ℃的区间, 高原鼠兔的洞口密度为69.01个· hm-2; 在陆地表面温度为30.0~30.5 ℃的区间内, 高原鼠兔的洞口密度为7.69个· hm-2(图6)。回归分析结果表明, 高原鼠兔的洞口密度随着陆地表面温度的增加而呈显著减少的趋势(y=-159.01x+4 799.7, R2=0.42, P< 0.05)(图6)。
 | 图6 高原鼠兔洞口密度在不同陆地表面温度区间的分布特征及其与陆地表面温度的关系Fig. 6 Plateau pika burrow densities in different land surface temperature (LST) interval and its relationship with LST |
青藏高原鼠兔通常构建庞大、复杂的洞穴系统来提高自身的存活率, 其种群密度与洞口密度呈显著正相关关系[15]。中国科学院西北高原生物研究所与青海省草原总站鼠害联合地面调查中, 使用总洞口数, 结合堵洞盗洞和夹捕法可以获得高原鼠兔有效洞口数和高原鼠兔种群密度[27]。因此, 高原鼠兔总洞口密度的分布格局可以有效反映其种群密度的情况。同时, 栖息地环境的变化往往会导致动物群落及其种群密度的变化, 在一定条件下, 栖息地环境要素的特征可能是影响小型哺乳动物分布和种群密度的重要因子[28]。本研究表明, 不同草地类型高原鼠兔的洞口密度存在显著差异(P< 0.05)。 在青海省达日县的研究也发现, 不同退化程度草地之间高原鼠兔种群数量差异显著[15]。本研究表明, 最大的高原鼠兔洞口密度出现在高寒草甸, 这与适度退化的草地植被高度较低、视野开阔, 可以为高原鼠兔提供理想的栖息地[14, 15]的观点并不一致。可能是因为以往的研究对于适度的退化草地的定义并没有统一的量化标准[16]。本研究也表明, 在荒漠、灌丛区域, 高原鼠兔洞口密度较低, 可能是因为荒漠中高原鼠兔喜食植物比率较低[15], 土壤含水量较少[29], 灌丛中郁闭度较高所致。由于资源有限, 无法在相对短的时间内开展大量的工作, 因而会存在一定的误差, 今后需要在一些典型样地开展高原鼠兔洞口季节动态变化监测, 从而评价大范围调查的误差。
NDVI是目前广泛使用的植被指数, 可以较好地反映像元内的植被类型、覆盖状态、植被生产力等信息[30]。林鑫等[31]在研究物种丰富度的大尺度地理格局及其成因时指出, NDVI对模型解释率的贡献最大, 可能是影响哺乳动物物种丰富度地理分异的主导因子。本研究发现, 不同NDVI区间里高原鼠兔的洞口密度存在显著差异(P< 0.05), 这与在四川省石渠县的研究结果[16]相一致。本研究表明, 高原鼠兔洞口密度在NDVI为0.5~0.6的区间里最高。这可能是因为NDVI值较高时, 往往表现为较高的植被高度, 从而使得草地环境的“ 郁闭度” 较高[15], 缩小了高原鼠兔的视觉范围, 降低了其逃生速度, 增加了潜在的捕食风险[14]。在野外调查中也发现, 高原鼠兔生性机警, 往往出洞活动前, 都要仔细观察周围环境, 确认没有危险才离开洞口。在NDVI为0.4~0.5时, 地上植被以灌丛为主, 高原鼠兔洞口密度最低。可能是因为灌丛的密度较大, 不利于其洞穴生活[29], 而且灌丛中高原鼠兔喜食植物比率较低[15]。当NDVI值低于0.2时, 高原鼠兔洞口密度较低, 说明高原鼠兔避免生存于退化程度严重的草地, 客观上防止了草地的过度退化。才让吉等[32]指出, 鼠类与家畜的食物资源谱存在较大差异, 高原鼠兔的食谱较为宽泛, 因食物竞争而对家畜构成的不利影响较为有限。由于青藏高原恶劣的自然环境和道路限制, 本研究所设置的航拍工作点并未涉及到NDVI值更高的藏东南地区; 同时由于技术手段限制, 目前也难以高效准确地获取地表群落特征。在下一步的工作中需要继续扩大航拍工作点, 获取更多的航拍照片, 利用软件准确获取地表群落特征, 以便更精确地评估NDVI对高原鼠兔洞口密度的影响。
土壤含水量不仅是反映土壤质量的核心指标之一, 同时也是影响植被分布的重要因素[33]。在一定程度上, 高原动物选择栖息地会受到土壤水分的影响[29, 34]。同时, 动物的挖掘行为也会显著改变土壤的入渗率, 降低地表径流[8]。在中国科学院海北站的研究中也发现, 高原鼠兔的活动可增加高寒草甸土壤表层含水量[9]。本研究显示, 不同土壤水分区间里高原鼠兔的洞口密度存在显著差异(P< 0.05), 高原鼠兔洞口密度在土壤含水量为20%~25%的区间里最高, 可能是因为土壤水分太高不利于高原鼠兔筑巢; 而土壤水分太低的区域, 地上植被以荒漠为主, 高原鼠兔喜食植物比率较低, 无法提供充足的食物[15]。但是由于青藏高原地形复杂、环境恶劣、观察站点分布稀疏, 本研究使用的水分数据分辨率为0.25° × 0.25° (约25 km× 25 km), 不能够精确地反映像元内部土壤水分的差异情况。因此, 在将来的研究需要空间尺度更高精度的水分数据, 以进一步探知土壤含水量对高原鼠兔洞口密度的影响。
陆地表面温度通常被视为影响植被状况的重要因素之一[5, 35]。已有许多研究利用陆地表面温度与NDVI的关系来探究植被生长的限制因子[36]。陆地表面温度是高寒草甸生长的主要限制因子[24], 而植被生长状况直接影响高原鼠兔的种群密度[37]。本研究显示, 不同地表温度区间里高原鼠兔的洞口密度存在显著差异(P< 0.05), 高原鼠兔洞口密度在生长季陆地表面温度为28.5~29 ℃的区间里最高, 可能是因为在该温度区间里, 地上植被以高寒草甸、草原化草甸、退化草甸、退化沼泽和沼泽为主, 相较于灌丛, 这些生境里的植物高度较低, 具有更低的“ 郁闭度” , 有利于高原鼠兔抵御高原上其它动物的捕食, 提高其生存几率, 相较于荒漠、草原, 这5种草地类型具有更高的NDVI值, 因而鼠兔洞口密度在该温度区间里最大。而陆地表面温度较高的区域, 地上植被主要以灌丛、荒漠为主, 高原鼠兔洞口密度最低, 有可能是因为灌丛密度大, 不利于高原鼠兔的洞穴生活[29]; 而荒漠中高原鼠兔喜食植物比率较低[15]、土壤含水量较少, 无法保证高原鼠兔充足的食物来源。
本研究采用无人机航拍和遥感数据相结合的方法初步探讨了高原鼠兔的空间分布及其栖息地的环境特征。结果发现, 高寒草甸高原鼠兔洞口密度显著高于其它草地类型; 此外, 高原鼠兔洞口密度最大值分别出现在NDVI值为0.5~0.6, 土壤含水量为20%~25%和生长季陆地表面温度为28.5~29 ℃的范围内。以上结果说明, 植被和土壤水热是影响高原鼠兔选择栖息地的主要因子, 高原鼠兔偏向于选择植被条件较好、土壤含水量较高, 生长季陆地表面温度相对适宜的区域。本研究采用的无人机可以在20 m的高度获取航拍照片, 进而利用笔者团队自主开发的航拍照片处理软件可以清楚地识别高原鼠兔洞口信息, 为在样地和区域尺度开展高原鼠兔研究提供了一种新的方法。
(责任编辑 武艳培)
The authors have declared that no competing interests exist.
| [1] |
|
| [2] |
|
| [3] |
|
| [4] |
|
| [5] |
|
| [6] |
|
| [7] |
|
| [8] |
|
| [9] |
|
| [10] |
|
| [11] |
|
| [12] |
|
| [13] |
|
| [14] |
|
| [15] |
|
| [16] |
|
| [17] |
|
| [18] |
|
| [19] |
|
| [20] |
|
| [21] |
|
| [22] |
|
| [23] |
|
| [24] |
|
| [25] |
|
| [26] |
|
| [27] |
|
| [28] |
|
| [29] |
|
| [30] |
|
| [31] |
|
| [32] |
|
| [33] |
|
| [34] |
|
| [35] |
|
| [36] |
|
| [37] |
|