



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
科尔沁沙地植物的生态及分布区型结构分析
[姜汇 , 高凯, 周立业, 田迅]
, 高凯, 周立业, 田迅]
, 高凯, 周立业, 田迅]
|
|


第一作者:姜汇(1995-),女,内蒙古莫力达瓦达斡尔族自治旗人,在读硕士生,研究方向为田间杂草生理生态。E-mail:[email protected]
为调查科尔沁沙地草原近年来植物多样性和植物生态特征,以反映该地区环境变化和植物生态适应性。本研究于2013年8月选取科尔沁沙地共56个样地,对样地植物按典型的科、属、种分类系统和株生长型、根生长型、Raunkiaer生活型、水分生态型及分布区型进行调查和分类,结果表明,科尔沁沙地具有典型的地面芽植物气候、中旱生生境条件、植物分布区系结构复杂的特征,共发现121种植物,隶属于31个科,6个株生长型,5个根生长型,6个生活型,5个水分生态类型和13个分布区型。其中,禾本科最多,占23.1%;菊科和豆科分别占22.3%和11.6%。株生长型结构以直立型种类最多,占37%;分枝型和丛生型分别占30%和19%。根生长型结构以直根型的种类最多,占61%;刷状根型和根茎型分别占22%和15%。生活型结构以地面芽植物种类最多,占35%;一年生植物和地面-地下芽植物分别占26%和14%。水分生态型以中生型植物种类最多,占44%;旱生型和中旱生型分别占26%和23%。分布区型结构中,蒙古-东北-兴安-华北分布区型(Mo-N-D-C)的植物种类占据首位,达30%;接下来是蒙古分布区型(Mo)和蒙古-东北-华北(Mo-N-C)分布区型,分别占18%和10%。结果表明,科尔沁沙地草原目前处于生态恢复的早期阶段,植物区系较复杂,水分环境没有发生重大变化,有利于该地区植物正向演替,但表层土壤生态环境依旧脆弱,一年生植物数量占绝对优势,应继续加强草地管理,合理保护和利用植物资源。
To investigate plant diversity and ecological characteristics of plants in Horqin sandy land, and to reflect environmental change and ecological adaption traits, 56 sample plots were established in 2013. Plants were classified according to growth form of the species, growth form of the roots, Raunkiaer life form, ecological type of water, and areal type. There were 121 species of plants in this region, belonging to 31 families. There were 6 plant growth forms, 5 root growth forms, 6 life forms, 5 ecological water types, and 13 areal types. Gramineae was the most abundant accounting for 23.1% of the plants, and composites and legumes accounted for 22.3% and 11.6%, respectively. Plant growth forms were primarily the vertical type, accounting for 37%, followed by the branch type and cluster type that accounted for 30% and 19%, respectively. Of the root growth forms, the taproot type was the most abundant, accounting for 61%, followed by brush root and rhizome type that accounted for 22% and 15%, respectively. The structure of life style was primarily hemicryptophyte species, accounting for 35%, and annual plants and ground geophytes accounted for 26% and 14%, respectively. The ecological water type was primarily the born in, accounting for 44%, followed by the xeric and mcsoxerophyte type that accounted for 26% and 23%, respectively. Distribution structure of the species was primarily the Mongolian-Northeastern-Dahuricia-North China, accounting for 30%, followed by the Mongolia area and Mongoian-Northeastern-North China areal type that accounted for 18% and 10%, respectively. These results indicate that the Horqin sandy land is at the early stage of ecological recovery. Its flora is complex, with a lack of large changes in water conditions, which are beneficial for the positive succession of plant communities. However, the top soil is still vulnerable, and most of plants are annual plant species. Thus, improvement of grassland management is still needed to protect and properly utilize the plant resources.
科尔沁沙地草原位于我国东北西部地区, 地处温带半干旱草原地带, 主体位于内蒙古赤峰市和通辽市内, 总面积约5.06万km2, 属温带大陆性季风气候, 植被多样性丰富[1]。近几十年来, 由于自然条件和人类活动的综合影响, 该草原生态环境发生了剧烈变化。20世纪60-90年代, 科尔沁沙地草原年均气温升高0.5~1.2 ℃, 沙漠化面积由32 925 km2增加到71 884 km2, 年均增加速度高达3.94%[2, 3]。物种多样性和均匀度下降, 生态优势度上升; 牧草产量和质量显著下降, 优良牧草数量减少, 适口性较差和有毒有害植物增加[4]。气候的暖干、生境干旱化、土壤盐渍化和过度放牧是造成科尔沁沙地草原退化的主要原因, 其中过度放牧是造成草甸植被退化的直接原因[4]。李爱敏等[5]在2000-2005年的研究发现, 科尔沁沙地荒漠化速度为每年0.14 km2, 表明该地区沙漠化速度减慢, 但沙漠化趋势并没有改变。
沙漠化是伴随物种多样性衰减的过程, 具体表现为首先导致特有物种的灭绝, 紧接着为稀有物种和普通物种的灭绝[6], 与之而来的是生物资源锐减和生态环境的进一步恶化[7]。面对科尔沁沙地草原日益严重的生态恶化问题, 有学者对恢复科尔沁沙地植被变化进行了相关研究, 常学礼和邬建国[6]早在1997年对科尔沁沙地沙漠化过程中的物种多样性的研究中发现, 多年生草本和灌木类植物受沙漠化影响最大, 而一年生草本和半灌木植物可在沙漠化过程的各个阶段存活, 在沙漠化过程中植物的灭绝速率大于定居速率。张继仪等[8]用围封的方法研究了科尔沁沙地在植被恢复过程中植物群落结构和物种多样性, 结果表明, 在演替初期一年生植物占绝对优势, 在演替早期藜科植物占有很高的优势度, 但随演替进行, 多年生植物的种类和数量逐渐超过一年生植物, 禾本科植物种类和优势度随演替进行不断增加, 群落生态优势度下降, 均匀度增加, 群落趋向稳定。对封育20年间固定沙丘的植被演替变化研究发现, 植被丰富度、盖度和物种多样性逐渐增加, 植被恢复演替模式为以先锋植物沙蓬(Agriophyllum squarrosum)为主的一年生植物群落→ 灌木差巴嘎蒿(Artemisia halcden)和一、二年生草本植物为主的群落→ 一、二年生草本和多年生草本为主的杂草群落[9]。在演替初期, 一年生草本植物在各群落中占绝对优势, 之后多年生植物种类明显增加, 藜科植物在演替早期有很高的优势度, 之后逐渐下降, 禾本科植物的种数和优势度逐渐增加, 在群落中的地位逐渐增强。整个演替过程中植物多样性增加, 地上生物量成倍增加, Simpson指数和Shannon-Wiener指数均呈增加趋势[10]。
以往对植被恢复过程中的研究主要围绕生物多样性、Simpson指数、Shannon-Wiener指数、物种优势度、丰富度以及植被盖度等为指标进行, 而少以植物的生态特征为依据研究科尔沁沙地植被分布情况。植物的生态特征是植物对一种或一组重要环境因子长期适应的结果, 是指植物在环境中表现出具有地带性特点的各种生理或生态特征, 对环境具有一定的指示作用[11, 12]。植物分类学中, 依据科、属、种等传统分类方法具有主导地位, 但并非是划分植物类别的唯一方法[12], 有学者依据植物的生态特征将其分成诸如生长型、Raunkiaer生活型、水分生态类型及分布区型等生态类型, 并建立了许多生态学分类系统[11, 13, 14]。
植物的生长型反映植物生活的环境条件, 相同的环境条件具有相似的生长型, 是植物趋同适应的结果, 生态环境的恶化必然导致植物群落生态特征的改变, 本研究根据实地考察, 在整个科尔沁沙地草原通过对56个样地植物进行系统记录和分类, 并依据植物生态特征对科尔沁沙地的植物组成进行了植株生长型、生活型、根生长型、水分生态类型及分布区型[15, 16]等结构分析, 以期了解科尔沁沙地草原近年来植物组成, 同时根据不同的生活型普反映该区域环境变化, 为科尔沁沙地植被恢复和相关研究提供基础资料。
试验样地位于科尔沁沙地草原, 该地区夏季炎热, 冬季干燥寒冷, 年平均降水量在350~450 mm, 且集中分布在6-8月, 雨热同期, 春秋以西南风为主, 冬季以西北风为主, 年平均风速为3.5 m· s-1, 最大风速可达21.7 m· s-1, 年日照2 900 h, 日照率67%, 无霜期为140~150 d, 为典型的半干旱大陆性季风气候[11], 该区域与东北、蒙古、兴安和华北4个植物区系相临。调查样地附近主要植被有羊草(Leymus chinensis)、糙隐子草(Cleistogenes squarrosa)、达乌里胡枝子(Lespedeza davurica)、差巴嘎蒿以及猪毛菜(Salsola collina)等。
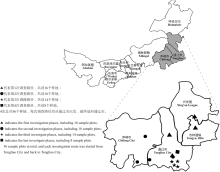
1.2.1 样地设置 从阿鲁科尔沁旗(天山)、扎鲁特旗、科尔沁右翼前旗(高力板)、八仙筒、新惠、林东、金宝屯、大青沟、甘旗卡、木里图、奈曼、开鲁、珠日河等旗县以及各旗县下属苏木、嘎查取样调查, 共设置56个样地, 取样地点如图1所示。样地主要代表植物有羊草、糙隐子草、达乌里胡枝子、差巴嘎蒿、猪毛菜等, 且以群落方式分布。
 | 图1 科尔沁沙地草原调查路径点示意图Fig. 1 Investigation places marked in the map with different signs in Horqin sandy land |
1.2.2 调查项目和方法 2013年8月, 在草原植物生长旺季进行野外实地调查。每个样地随机置4~8个50 cm× 50 cm的样方, 分别对每个样方内的所有植物逐一按株生长型、根生长型、Raunkiaer生活型、水分生态类型和分布区型进行统计。参照杨允菲和祝廷成[17]的分类办法, 依据植株地上形态将株生长型分为直立型、莲座型、半莲座型、丛生、分枝和匍匐, 依据根的形态特征将根生长型分为刷状根、直根、块根、块茎、鳞茎和鲜茎, 依据休眠或复苏芽所处的位置高低和保护方式将Raunkiae生活型分为高位芽植物、地上芽植物、地面芽植物、地面-地下芽植物、地下芽植物和一年生植物。水分生态型分为旱生、中旱生、中生、中湿生、湿生、水生, 分布区型分为东北分布区型、华北分布区型、蒙古分布区型、兴安分布区型、蒙古-东北-兴安分布区型、蒙古-东北分布区型和蒙古-东北-兴安-华北分布区型。按上述标准对各地区每个样方内出现的所有植物按科、属、种进行分类并记录, 然后对记录的所有植物按上述分类标准分类, 并统计各分类系统的数量及百分比, 将作为其结构分析的指标[18]。
通过对科尔沁沙地56个样地的调查发现共有121种植物(表1), 分布于31个科。其中禾本科植物种类最多, 为28种, 占23.1%; 其次为菊科, 27种, 占22.3%; 豆科14种, 占11.6%; 藜科8种, 占6.6%; 唇形科6种, 占5.0%; 蔷薇科3种, 占2.5%; 百合科、毛茛、大戟科、旋花科、蓼科、苋科、石竹科、紫草科、鸢尾科和十字花科10个科均分别为2种, 各占1.7%; 莎草科、瑞香科、车前科、马齿苋科、萝摩科、麻黄科、蒺藜科、亚麻科、桑科、牻牛儿苗科、紫葳科、胡颓子科、蝶形花科、伞形科和榆科15个科均分别为1种, 各占0.8%。
| 表1 科尔沁沙地植物的种类及生态组成 Table 1 Plant species and their ecological components in Horqin sandy land |
植物的生长型及生活型都是植物与环境长期的协同进化所形成的形态和外貌特征。按株生长型分类, 科尔沁沙地植物以直立型的种类最多, 为45种, 占37%; 其次是分枝型, 为36种, 占30%; 第3是丛生型, 为23种, 占19%; 匍匐型为8种, 占7%; 半莲座型为5种, 占4%; 莲座型为4种, 占3%(图2)。按根生长型分类, 科尔沁沙地直根型植物最多, 为74种, 占61%; 其次为刷状根型, 27种, 占22%; 根茎型植物有18种, 占15%; 鲜茎型和鳞茎型各1种, 分别仅占1%(图3)。按Raunkiaer对休眠芽位置的生活型分类来看, 地面芽植物种类最多, 为42种, 占35%; 一年生植物次之, 为32种, 占26%; 地面-地下芽植物位居第3, 有17种, 占14%; 地下芽植物有15种, 占12%; 地上芽植物有8种, 占7%; 高位芽植物最少, 仅7种, 占6%(图4)。
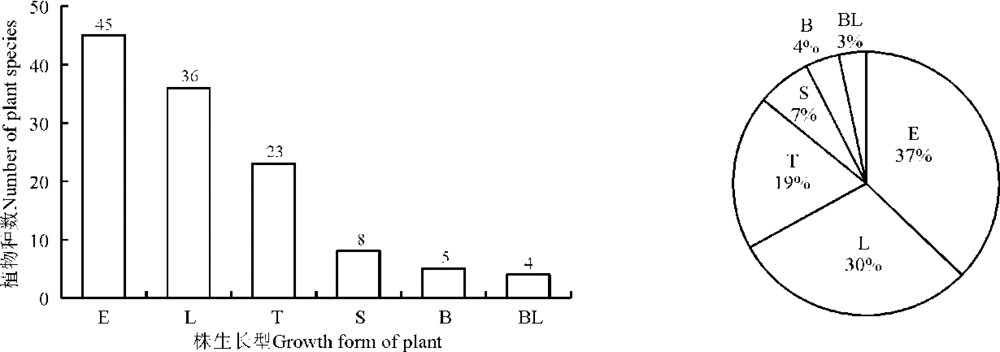 | 图2 科尔沁沙地植物株生长型的数量分布及株生长型谱Fig. 2 Quantitative distribution and spectrum of growth form of plant species in Horqin sandy land |
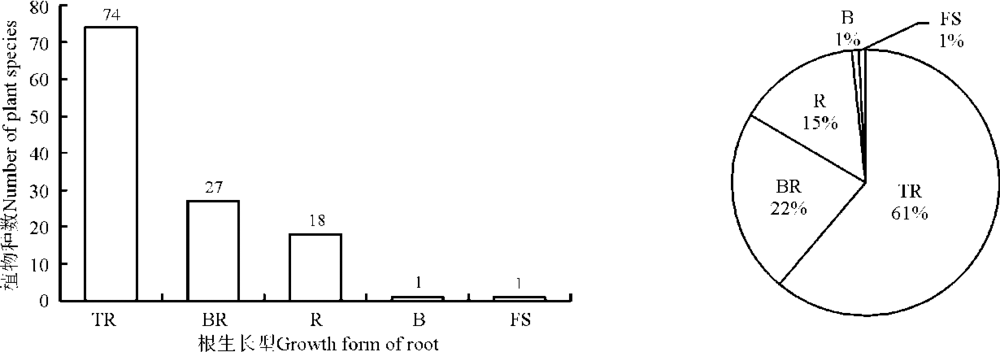 | 图3 科尔沁沙地植物根生长型的数量分布及根生长型谱Fig. 3 Quantitative distribution and spectrum growth form of roots on plant species of Horqin sandy land |
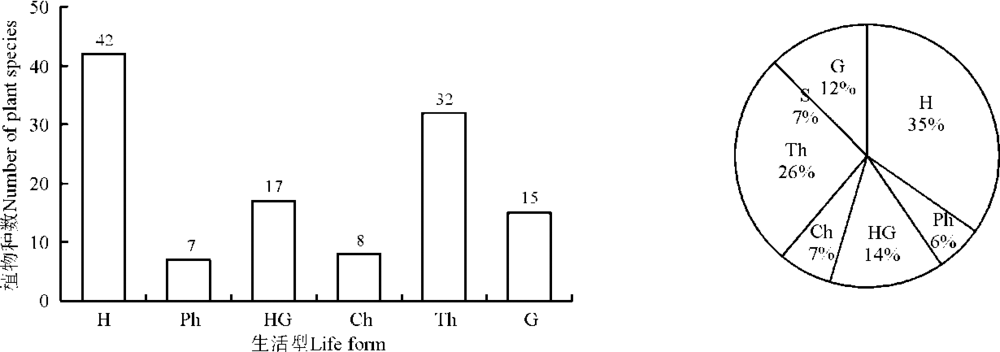 | 图4 科尔沁沙地植物生活型的数量分布及生活型谱Fig. 4 Quantitative distribution and spectrum of life form on plants species of Horqin sandy land |
科尔沁沙地植物在水分生态类型的数量分布中, 中生型植物最多, 为54种, 占植物种类总数的44%, 具有明显优势; 旱生型和中旱生型植物次之, 各为31种和28种, 分别占植物种类总数的26%和23%; 而湿中生类型和湿生型植物数量稀少, 各为6种和2种, 共占7% (图5)。由此反映出科尔沁沙地土壤的水分条件为中生-干旱类型, 干旱趋势比较明显。
 | 图5 科尔沁沙地植物水分生态类型的数量分布及水分生态类型谱Fig.5 Quantitative distribution and spectrum of ecological type of water on plants species of Horqin sandy land |
由于不同植物具有不同的广布性, 有时可能同一植物同时隶属于好几个地理成分[11]。科尔沁沙地植物共有13种分布区型(图6), 其中以蒙古-东北-兴安-华北分布区型(Mo-N-D-C)数量最多, 为37种, 占植物种类总数的30%; 蒙古分布区型(Mo)次之, 为22种, 占植物种类总数的18%; 蒙古-东北-华北分布区型(Mo-N-C)、蒙古-华北分布区型(Mo-C)、蒙古-兴安(Mo-D)和东北-华北分布区型(N-C)为第三梯队, 各为17种、12种、10种和10种, 分别占植物总体的14%、10%、8%和8%; 华北分布区型(C)、兴安分布区型(D)、蒙古-东北(Mo-N)、兴安-华北分布区型(D-C)、蒙古-东北-华北分布区型(Mo-NC-)、东北分布区型(N)以及东北-兴安-华北分布区型(N-D-C)少量存在于科尔沁沙地中, 数量分别在1~3种不等, 共占植物种类总数的12%。由此可以看出, 科尔沁沙地植物的分布区型比较混杂, 属蒙古、东北、兴安、华北四系植物种类复杂交汇地域。
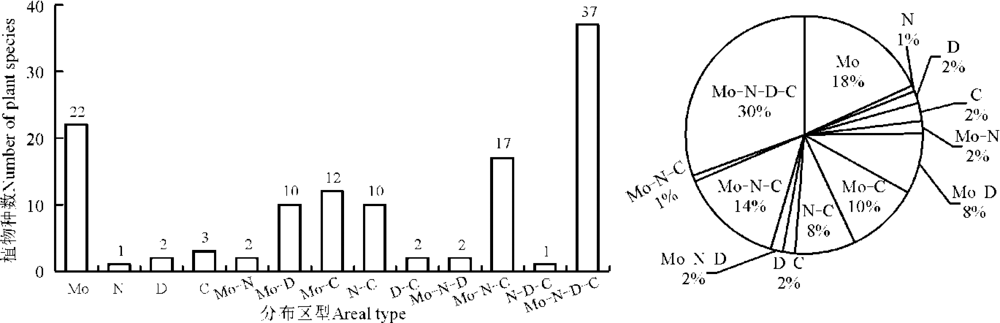 | 图6 科尔沁沙地植物分布区型的数量分布及分布区型谱Fig.6 Quantitative distribution and spectrum of areal type on plants species of Horqin sandy land |
研究表明, 放牧是造成科尔沁沙地退化的直接原因, 封育和禁牧可有效改善科尔沁沙地草原植物群落, 有利于该地区植被恢复[19]。在国家禁牧政策和草原补偿机制实施的大背景下, 2003年内蒙古自治区开始全面禁牧, 并实施草原生态补偿机制[20], 自此该地区草地群落得到有效保护, 植被逐渐恢复[21]。
植物群落中的物种组成及其优势种的分布特征基本决定了植物群落外貌与结构类型的基本特征[22]。在温带草原区, 禾本科和豆科植物大多数物种均为优良牧草, 菊科和藜科植物大多数物种为中、劣质杂草, 而毛茛科和紫草科植物大多数物种有毒或有害, 本研究发现科尔沁沙地中禾本科、菊科和豆科植物种类分别位居第1、2和第3位, 禾本科和豆科植物种类排名靠前, 表明其饲用价值较高, 在施行适当保护措施的前提下可以进行放牧或刈割, 另外, 也发现毛茛科和紫草科植物各两种。研究表明, 在演替早期, 黎科植物占有很高的优势度[8, 10], 而本研究发现黎科植物仅7种, 占5.8%, 禾本科和豆科占有优势, 表明科尔沁沙地处于群落演替的早期向中期进行。武晶等[23]在2011年对科尔沁沙地调查的研究中发现共有83种植物, 隶属于19科21属; 本研究比武晶等[23]的调查多出28种12科7属, 可能原因:1)调查样地选择大小和路径不同, 2)2011-2013年两年内该地区植物种类和群落多样性随演替进行而增加。常学礼和邬建国[26]对科尔沁沙地荒漠化过程的研究表明, 多年生草本、木本和灌木类植物受荒漠化的影响最大, 是抵御荒漠化的重要植物; 一年生植物很大程度上受降水量及降水时机的影响, 在防治沙化过程中具有较大随机效应, 而本研究发现一年生植物种类占26%, 反映出科尔沁沙地草原生态系统较为脆弱的局面依然没有改变。
植物生长型是根据植物的可见结构分成的不同类群, 植物的生长型反映植物生活的环境条件, 相同的环境条件具有相似的生长型, 是趋同适应的结果。植物根的生长型可以反映植物的固着、吸收营养和水分, 甚至是营养繁殖能力。一般情况下刷状根对表层土壤的固着能力较强, 直根对深层土壤的固着能力较强, 根茎不仅对表层土壤的固着能力强, 而且具有较强的营养繁殖能力[24]。科尔沁沙地草原植物以直根为主, 占61%, 而刷状根和根茎植物共占37%, 说明当前该地区植物以固着深层土壤的植物为主, 而对表层土壤的固着能力相对较弱, 表层土壤稳定性比较脆弱; 根茎型植物占15%, 说明该地区繁殖能力较强的植物开始出现, 有利于该地区植被恢复。植被组成的Raunkiaer生活型谱可以反映群落的稳定性、局部环境条件状况以及发展方向。一年生植物以种子度过不良季节, 对恶劣环境有最大的适应性, 但其萌发及幼苗对环境的要求较严格, 在群落中具有较高的不稳定性。因此, 在Raunkiaer生活型中, 一年生植物的比例越大, 群落越不稳定[24]。本研究发现, 科尔沁沙地多为草本植物, 灌木、乔木相对贫乏, 且以地面芽植物和一年生植物为主, 多年生植物和高位芽、地上芽植物较少, 表明科尔沁沙地草原植物群落存在较高的不稳定性, 脆弱的生态局面依然没有改变, 需要继续采取保护措施以进行正向演替。当放牧降低了草原植物群落多年生植物功能群的生物量时, 一年生植物功能群在湿润季节或年份可补偿群落生物量或生产力[25], 科尔沁沙地草原高位芽和地上芽植物的出现也表明该地区环境在禁牧及相关保护措施下出现一定好转。
植物群落的形成是组成物种与外部环境相互作用的结果[26, 27], 土壤和水分是影响植物生长、发育的重要生态因子, 同时水分也是土壤最重要的组成部分[28, 29], 植物在不同的土壤和水分的生态适应中形成不同的水分生态型[30], 科尔沁沙地属于温带干旱半干旱气候区, 土壤含水量较低, 且与温度和日照时数呈负相关关系, 与降水量、相对湿度、蒸散量呈正相关关系[31, 32]。按水分生态类型来看, 科尔沁沙地草原植物中, 中生、旱生和中旱生植物分别位居1、2、3位, 说明该地区群落结构和功能主要由中生、旱生和中旱生植物决定, 而中生植物所起作用最大, 这与科尔沁沙地所处地区气候类型一致, 说明该地区气候类型没有发生重大改变。杨利民等[33]对我国东北羊草群落植物水分生态类型功能群的研究表明, 中旱生植物生物量与年降水量、土壤含水量、年均温呈正相关关系, 而旱生植物仅与年均温度因子呈峪形函数关系, 反映了不同植物水分生态类型对环境变化响应的敏感性不同, 中旱生植物对环境更为敏感, 其群落结构的稳定性对水分依赖性更高。刘江华等[34]在黄土高原对刺槐(Robinia pseudoacacia)林下植被的水分生态型和生活型的研究发现, 旱生、中旱生植物在群落中的重要值随林地年龄逐渐降低, 而中生生植物在群落中的地位逐渐提高, 林下植被由旱生向中旱生型转变。而本研究发现, 中旱生植物和旱生植物共占有70%的比例, 对环境较为敏感的中旱生植物也占有23%的比例, 因此从植物水分生态类型数据结构中可以得知:1)现时科尔沁沙地土壤水分类型为中旱生类型, 2)干旱化比较明显, 3)目前水分条件有利于植物演替。
科尔沁沙地草原是蒙古植物区系、东北植物区系、兴安植物区系、华北植物区系 4个植物区系相汇集的区域[35], 植物区系成分的生态类型结构和分布区型结构较为复杂, 表现为:1)一种植物同时属于不同的区系成分, 2)具有4个植物区系相汇集的区域植被特点。按植物区系成分来看, 有102种植物属于蒙古植物区系成分, 70种植物属于东北植物区系成分, 54种植物属于兴安植物区系成分, 82种植物属于华北植物区系成分, 未发现特有种。
本研究表明, 科尔沁沙地共有6个株生长型、5个根生长型、6个Raunkiaer生活型, 5个水分生态类型和13个分布区型, 总体来来看科尔沁沙地目前处于植物恢复的早期, 典型特征是禾本科植物和豆科植物数量较多, 一年生植物数量占绝对优势, 但水分环境没有发生重大改变, 有利于植被正向演替进行, 植物种类逐渐增加。应当继续采取封育或禁牧等保护措施以利于当地植物群落进行演替和恢复。白美兰等[21]的研究也表明, 科尔沁沙地荒漠化整体出现好转, 但在目前的气候条件下, 局部沙漠化有所恶化, 应当加强草地管理, 减少人为破坏或鼠害等[36], 合理保护和利用该区域植物资源, 为科尔沁沙地植被的逐步恢复做好保障。
(责任编辑 武艳培)
The authors have declared that no competing interests exist.
| [1] |
|
| [2] |
|
| [3] |
|
| [4] |
|
| [5] |
|
| [6] |
|
| [7] |
|
| [8] |
|
| [9] |
|
| [10] |
|
| [11] |
|
| [12] |
|
| [13] |
|
| [14] |
|
| [15] |
|
| [16] |
|
| [17] |
|
| [18] |
|
| [19] |
|
| [20] |
|
| [21] |
|
| [22] |
|
| [23] |
|
| [24] |
|
| [25] |
|
| [26] |
|
| [27] |
|
| [28] |
|
| [29] |
|
| [30] |
|
| [31] |
|
| [32] |
|
| [33] |
|
| [34] |
|
| [35] |
|
| [36] |
|