





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
玉米/大豆套作体系中大豆茎秆生长过程模拟
[杜勇利1  , 周涛
, 周涛1 , 杨欢1 , 刘婷1 , 官思成1 , 邓瑜川2 , 刘卫国1 , 杨文钰1 ]
, 周涛]
|
|


第一作者:杜勇利(1991-),男,四川宜宾人,在读硕士生,主要从事作物高产稳产优质高效栽培研究。E-mail:[email protected]
为建立套作系统中大豆( Glycine max)茎秆生长过程的模拟方程,本研究以玉米( Zea mays)/大豆套作为对象,单作大豆为对照,3种不同株型大豆(南豆032-4、南豆12、九月黄)为材料,对两种种植模式下大豆茎秆生长进行定量化模拟。运用Richards方程[ y=α/(1+βe- γx)1/δ],构建大豆茎长、茎粗生长变化的动态模型,方程参数α值(终极生长量参数)为各品种遗传参数,β值(初值生长量参数)、γ值(生长速率参数)和δ值(形状参数)在不同栽培模式、品种间变异较大, 可见主要通过调节参数β、γ、δ值来实现对整个方程的调控。采用Logistic方程[ y=α/(1+βe- γx )],构建大豆茎秆干重动态模型,β、γ值在不同栽培模式、品种间变异较大,主要通过调节参数β、γ值来实现对整个方程的调控。因此分别建立套作和单作栽培模式下的大豆模拟方程,构建大豆茎秆长、茎秆粗的生长动态模型,以及构建大豆茎秆干重积累的动态模型,所建立套作和单作生长模型方程精确度( R2)均在0.99以上。检验结果表明,模型对不同类型大豆材料在单套作体系下茎秆节间长度、粗度、干重都能很好的预测,能较好地拟合茎秆生长动态。本研究结果对探究玉米/大豆套作体系下大豆茎秆生长规律研究,具有一定的生物学意义。
, Zhou Tao
The intercropping of maize and soybean is practiced in several parts of the world. Using models for forecasting soybean growth is a good cropping technique; however, few research models are available that predict soybean growth in monocropping and intercropping practices. This study was based on the systematic observations of Nandou 032-4, Jiuyuehuang, and Nandou 12. The growth of soybean stems can be quantitatively simulated by the patterns of growth in monocropping and intercropping cultivation modes. A dynamic model was established using Richards equation for model building, where the time step was represented by the physiological development time (PDT), describing the elongation of soybean stem internodes and the enlargement of the dynamic processes at the internodes, in monocropping and intercropping practices. In the dynamic model, the time step was estimated by the growth and development taking place on the actual number of days by logistic equations, for predicting the dry weight of the soybean stem in monocropping and intercropping cultivation practices. From the model, the potential length and thickness of the internodes gradually increased as the number of soybean internodes increased and reached a maximum at the ninth and seventh internodes, respectively, and then gradually decreased. Therefore, the potential length of the ninth internode and the potential thickness of the seventh internode were identified as the potential genetic parameters that can be used to calculate the potential length and thickness of other internodes. At the same time, the potential dry weight of the stem was used to determine the potential dry weight of other stems. The inspection results showed that different varieties of soybean have different cultivation patterns. The model reflected a satisfactory prediction rate. This model, developed using soybean varieties having different modes of cultivation, internode lengths, internode thicknesses, physiological development times, and dry weights of the stem, has good predictability and reliability, and the stem growth dynamics predicted by the model conformed well to the biological rules of stem growth, highlighting the biological significance of the model.
大豆(Glycine max)起源于中国, 种植历史可以追溯到3 000年以前, 除了作为重要的粮油作物外, 其生物固氮、土壤磷活化、碳截获等生态意义越来越受到重视。以我国西南地区为例, 土壤发育程度浅, 有机质、速效氮、磷的含量低。但是受制于人口和粮食生产压力, 该地区复种指数高, 在旱地生态系统中, 间/套作占据主导地位[1]。套作豆科植物相比单作能显著提高产量, 主要体现在豆科植物活化土壤中难溶态磷、生物固氮供自身和共生作物吸收利用[2, 3, 4]。玉米(Zea mays)-大豆带状复合种植体系以其高产、生态等特点在我国西南地区得到大面积推广[5]。玉米/大豆套作模式不仅能使土壤水分、养分和光资源的供给得到改善, 还可以增加这些资源的可利用性, 产生互利, 从而提高复种系数[6]。间、套作种植模式不仅遵循了时间镶嵌原则、空间扩展原则、种间搭配原则和物能流畅原则, 而且还能解决用地和养地的矛盾、复种和季节不够用的矛盾, 同时, 作物茎秆还可解决发展畜牧业饲料不足的矛盾。
作物形态建成模拟是作物生长模拟中重要的一部分。明确作物外部形态变化规律, 定量化研究作物形态建成过程, 对指导作物生产具有重要的理论意义和实际应用价值。同时, 也是虚拟作物和作物可视化的研究基础[7]。我国西南地区属于寡日照区域, 在玉米和大豆共生期, 受高位作物玉米遮阴导致苗期大豆受到一定程度的阴蔽胁迫, 大豆植株茎长增加, 茎粗、光合速率和地上部生物量显著下降, 导致大豆易发生倒伏, 从而影响大豆产量[8, 9]。但是不同耐阴性大豆品种在套作环境中的茎秆生长特性呈现较大差异, 强耐阴大豆品种茎秆抗倒伏能力明显强于一般或不耐阴品种[10]。因此, 研究套作大豆的茎秆生长特性及不同品种茎秆在特殊光环境中的变化对于保障套作大豆产量和整个套作体系的稳产具有重要意义。
已有研究报道表明, 以生理发育时间(physiological development time, PDT)为步长, 可以对许多作物的形态生长动态进行模拟[11, 12, 13]。如, 不同施氮水平和水分条件下水稻(Oryza sativa)的地上部器官形态特征建成[14, 15]; 不同播期条件下大麦(Hordeum vulgare)茎秆、穗的生长动态[16]; 不同大豆品种茎、叶、荚的形态建成[17]。以生长日(growing degree days, GDD)为步长, Logistic方程可以很好的描述小麦叶片、穗、叶鞘和节间的动态生长变化[18, 19]。同样, 以扩充的Logistic模型可以对小麦(Triticum aestivum)株高和叶面积变化动态进行模拟[20]。此外, 利用系统分析方法和动态建模技术, 成功构建小麦叶片茎鞘夹角的动态模拟模型[21]。已有研究从不同角度对不同作物的生长发育规律进行了研究和模拟, 通过模型模拟植物的生长动态技术得到广泛应用。虽然对单作大豆的形态建成动态模拟已有研究, 但是套作大豆的生长环境与单作有较大差异, 所以已有模型不能完全适用于描述套作大豆茎秆的生长动态。
因此, 本研究以玉米/大豆带状套作复合种植模式中大豆为研究对象, 通过对大豆茎秆生长过程的系统观测与分析, 在定量研究的基础上, 分别构建套作和单作条件下大豆节间伸长和增粗生长的动态模型, 以及大豆茎秆干重变化动态模拟模型, 旨为构建大豆在不同种植模式下生长决策支持系统奠定基础, 也为实现大豆植株形态的虚拟显示、可视化研究提供关键技术。
3个供试大豆品种株型间差异较大, 分别为“ 南豆032-4” (无限型, 弱耐阴性, 四川省南充市农业科学院育种材料), 株高120 cm左右, 茎秆较细; “ 九月黄” (有限型, 中度耐阴性, 四川省阆中市地方品种), 株高75 cm左右, 茎秆粗细中等; “ 南豆12” (亚有限型, 强耐阴性, 四川省南充市农业科学院选育, 西南地区套作大豆主推品种), 株高90 cm左右, 茎秆较粗。玉米选用半紧凑型品种“ 川单418” (四川农业大学玉米所选育)。
试验于2012年和2013年在四川农业大学雅安校区教学科研园区(29° 98' N, 103° 00' E)进行, 属亚热带湿润季风气候区。年均温16.2 ℃, 年日照时数1 005~1 200 h, 年降水量1 250~1 750 mm; 土壤类型为紫色土, 质地为重壤土。pH 7.4, 有机质含量为27.86 g· kg-1, 全氮含量为1.79 g· kg-1, 全磷含量为0.89 g· kg-1, 全钾含量为29.76 g· kg-1, 速效氮含量为115 mg· kg-1, 速效磷含量为37 mg· kg-1, 速效钾含量为136 mg· kg-1。田间采用随机区组排列, 每个区组内包含3个玉米/大豆套作处理(3个大豆品种)和3个大豆单作处理, 小区面积24 m2, 重复3次。套作模式采用宽窄行种植, 玉米窄行行距40 cm, 宽行行距160 cm, 穴距35 cm, 每穴2株, 分别于当年4月初播种玉米。大豆于玉米宽行间播种两行, 窄行行距为50 cm, 穴距10 cm, 穴留2株, 玉米/大豆间距55 cm, 分别于2012年6月22日和2013年6月23日播种。单作模式中, 大豆行距50 cm, 穴距10 cm, 每穴定苗1株。氮(N)、磷(P2O5)、钾(K2O)用量分别为195、75和105 kg· hm-2; 玉米于每年4月初挖窝点播, 窝深7 cm左右, 宽15 cm左右, 将30%的氮和全部的磷、钾肥施于窝的一侧, 另一侧放种子避免种子与肥料接触, 而后覆土, 种子覆土深度不超过5 cm; 再于玉米拔节期和大喇叭口期分别追施30%、40%的氮, 均采用兑清水冲施于株旁的方式。大豆不施肥, 田间杂草病虫害管理方式同当地农户高产田。
由于大豆第1节和第2节的叶片分别为子叶、真叶, 从第3节间才开始长出复叶。因前两个节均不会开花结荚, 且节间长度较短, 同时品种间差异不大, 所以本研究中所有指标的模拟值和观测值均从第3节间开始。从大豆第3节始, 每小区选长势一致的大豆植株挂牌标记, 测定以下指标。
1.3.1 茎秆生长的观测 每隔3 d定点定株测量已标记大豆茎秆第3节及其以上各节的节间长度和粗度。节间长度使用米尺测量, 粗度使用游标卡尺测量, 其测量部位为节间中部。
1.3.2 茎干重测定 自播种大豆后7 d (即套作和单作大豆生育期都为VE)开始调查取样。每小区选取生长一致的300株挂牌标记。每隔7 d取样, 直至大豆收获。每次选取10株从茎秆第1节剪断, 单株按叶片、茎分解植株, 然后分别装入牛皮纸袋中, 置于烘箱中105 ℃杀青30 min, 再于75 ℃下烘干至恒重, 最后称重, 计算干重。
1.3.3 数据处理及建模 本研究以生理发育日(PDT)为步长, 运用Richards方程构建大豆茎长、茎粗生长的动态模型, 获取方程特征参数α 、β 、γ 、δ , 参数α 为增长量的极限值、β 为初始值参数、γ 为增长速率参数、δ 为曲线形状参数。方程模拟图形是以α 为渐近线的S形曲线。以作物生长发育实际天数为步长, 运用Logistic方程构建大豆茎干重积累的动态模型, 获取方程特征参数α 、β 、γ , 参数α 为增长量的极限值, β 、γ 均为方程特征参数。利用 Curve Expert 1.3 软件对单作和套作大豆的生长动态进行拟合, 探明生理发育时间与大豆茎秆生长的关系, 建立大豆生长动态模型所需的量化关系。大豆各发育阶段的生理发育时间的计算参照陈传梅等使用的方法[22]。并利用SPSS 17.0 软件分析这些特征参数。2012年试验数据用于模型构建和参数调试, 2013年数据用于模型检验。采用根均方差RMSE来对模型进行检验, 统计分析模拟值与观测值之间的符合情况。RMSE值越小, 说明模拟值与观测值的一致性越好, 模型的模拟结果越精确、可靠。RMSE的计算公式如下:
RMSE=
式中:OBSi为观测值, SIMi为模拟值, n为样本容量。优化不同大豆品种在不同栽培模式下, 建立具有生物学意义的动态共性模拟方程, 同时使用Excel分析软件进行回归分析。
3个供试大豆品种的茎秆节间潜在长度、茎秆潜在粗度和茎秆潜在干重如表1所列。3个供试大豆品种的遗传参数, 品种间和不同种植模式下都表现出了差异。且在套作种植模式下品种间的节间潜在长度、潜在粗度和茎的潜在干重均表现出显著差异性; 单作种植模式下, 品种间的遗传参数也表现出差异, 说明不同种植模式会改变品种的遗传参数。
| 表1 单作、套作下的大豆节间潜在长度、潜在粗度和茎的潜在干重 Table 1 Genetic coefficients of the three varieties of soybean |
2.2.1 大豆茎秆节间伸长的动态模型 试验结果表明, 在套作模式下大豆各节间从开始伸长到定长需要16.53个生理日, 各节间开始伸长的时间间隔为2.8个生理日。大豆第3节间开始伸长的生理发育时间为11.21个生理日。在单作模式下大豆各节间从开始伸长到定长需要14.88个生理日, 各节间开始伸长的时间间隔为1.9个生理日。大豆第3节间开始伸长的生理发育时间为8.95个生理日。各节间伸长的动态方程分别如下: 个大豆品种在套作和单作栽培模式下的方程, 其相关系数均在0.99以上。α 为每个品种潜在极限值, β 值和γ 值在栽培模式和不同大豆品种间变异均较大。δ 值在单作栽培模式下变异较小, 套作模式下变异较大。因此, 需要通过对套作和单作栽培模式下的大豆分别建立共性模拟方程。
套作:Ln(PDT)=
单作:Ln(PDT)=
式中:Ln(PDT)为大豆在某一生理发育时刻的第n节的节间长度(cm); Lmaxn为大豆第n节节间潜在长度(cm)。α 、β 、γ 、δ 均为模型参数(表2)。可以看出, 套作和单作栽培模式下, α 、β 、γ 、δ 值变异均大。但是, 3
| 表2 单作、套作下不同大豆品种相对节间长度的方程系数 Table 2 Coefficients of normalized internode lengths calculated using the dynamic equations in three varieties of soybean in intercropping and monocropping practices |
套作模式下, 随着大豆节间数的增加, 其各节节间潜在长度Lmaxn逐渐增长, 均在第7节间达到最大值, 然后又逐渐下降(图1)。因此, 在套作栽培模式下对南豆032-4、南豆12和九月黄的前7节茎秆节间长与PDT进行共性化处理, 得到如下方程:
Lmaxn=
单作模式下, 随着大豆节间数的增加, 其各节节间潜在长度(Lmaxn)逐渐增长, 不同品种大豆均在第12节间达到最大值, 然后又逐渐下降(图1)。因此, 在单作栽培模式下对南豆032-4、南豆12和九月黄的前12节茎秆节间长与PDT进行共性化处理, 得到如下方程:
Lmaxn=
2.2.2 节间增粗的动态模型 大豆节间增粗是从慢到快再到慢的过程。在套作模式下, 大豆各节间从开始增粗到粗度固定需要14.88个生理日, 各节间开始增粗的时间间隔为1.9个生理日。在单作模式下, 大豆各节间从开始增粗到粗度固定需要16.53个生理日, 各节间开始增粗的时间间隔为2.8个生理日。各节间增粗的动态方程如下:
套作:Ln(PDT)=
单作:Ln(PDT)=
方程式中:Ln(PDT)为大豆在某一生理发育时刻的第n节的节间粗度(cm); PDT为生理发育时间, 8.95为在套作中大豆第3节间开始增粗的生理发育时间, 11.21为在单作中大豆第3节间开始增粗的生理发育时间。Lmaxn为大豆第n节节间的潜在粗度, 是品种遗传参数。
可以看出(表3), 3个大豆品种在不同栽培模式下的方程, 其相关系数均在0.94以上, α 为潜在极限值; β 值和γ 值在栽培模式和不同大豆品种间变异均较大。δ 值在单作和套作栽培模式下变异均较大。
 | 图1 单作、套作下各节间潜在长度随节间数的动态变化Fig. 1 Changes in internode length with respect to the number of internodes |
| 表3 单作、套作下不同大豆品种相对节间粗度的方程系数 Table 3 Coefficients of normalized internode thickness calculated using the dynamic equations in three varieties of soybean in intercropping and monocropping cultivation practices |
套作模式下, 随着大豆节间数的增加, 其各节节间潜在粗度Lmaxn逐渐增加, 在第5节间达到最大值, 然后又逐渐下降(图2)。因此, 在套作栽培模式下对南豆032-4、南豆12和九月黄的前5节茎秆节间长与PDT进行共性化处理, 得到如下方程:
Lmaxn=
单作模式, 随着大豆节间数的增加, 其各节节间潜在粗度INLmaxn逐渐增加, 在第7节间达到最大值, 然后又逐渐下降(图2)。因此, 在单作栽培模式下对南豆032-4、南豆12和九月黄的前7节茎秆节间长与PDT进行共性化处理, 得到如下方程:
Lmaxn=
2.2.3 大豆干物质量动态变化模型 大豆地上生物量干质量呈“ 缓慢生长-快速生长-缓慢生长” 的生长过程。开始生长时较为缓慢, 随着时间的推移(或温度的升高), 在某一段时间内增长速度很快, 当达到某一值后, 增长速度又趋于缓慢, 直至最后停止生长。这一特点符合 Logistic生长曲线。利用 Curve Expert 1.3软件对单作和套作大豆茎干重和生长发育实际天数之间的关系进行拟合, 得到最优化的Logistic方程y=α /(1+β e-γ x)], 其中, y为干重, x为实际生长天数, 获得方程特征参数α 、β 、γ (表4)。
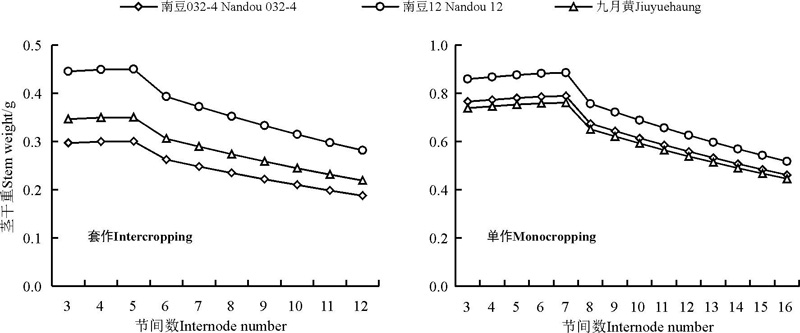 | 图2 单作、套作下各节间潜在粗度随节间数的动态变化Fig. 2 Changes in internode thickness with respect to the number of internodes |
| 表4 单作、套作下不同大豆品种相对茎秆干重的方程系数 Table 4 Coefficients of normalized dry weights of the stems calculated by dynamic equations in three soybean varieties in intercropping and monocropping cultivation practices |
在不同栽培模式下对南豆032-4、南豆12和九月黄的茎秆干重与实际生长天数进行共性化处理。
套作种植模式下的方程:
Wn(PDT)=
式中:Wn(PDT)为套作大豆在某一实际生长时刻的茎干重(g); n为生长实际时间(d); 通过动态观测套作大豆结果得出, Wmax77为大豆在第77天的时候大豆茎潜在干重达到最大(图3)。
单作种植模式下的方程:
Wn(PDT)=
式中:Wn(PDT)为单作大豆在某一实际生长时刻的茎干重(g); n为生长实际时间(d); 通过动态观测单作大豆得出, Wmax75为大豆在第75天的时候大豆茎潜在干重达到最大(图3)。
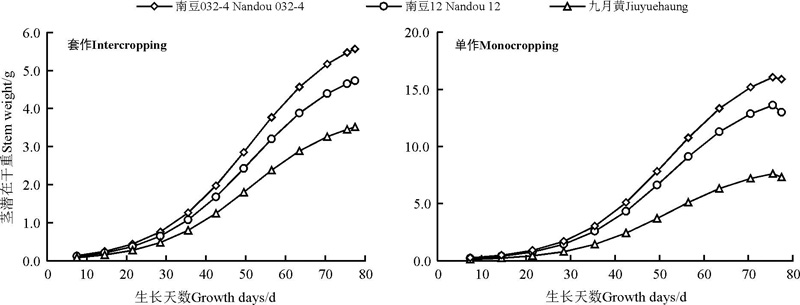 | 图3 大豆茎潜在干重随生长时间的动态变化Fig. 3 Dynamic changes of potential stem dry weight of intercropping and monocropping soybean |
通过2012-2013年的试验数据对套作和单作大豆茎秆动态模型进行检验。不同栽培模式下不同品种的大豆茎秆模拟值的RMSE结果(表5)表明, 套作大豆节间长度、粗度、干重的模拟误差分别为0.24~0.33 cm、0.007~0.030 cm、0.21~0.37 g。单作大豆节间长度、粗度、干重的的模拟误差分别为0.11~0.17 cm、0.021~0.027 cm、0.46~0.54 g。说明套作模式下的模型和单作模式下的模型模拟值与观测值都表现出很好的一致性, 模型的模拟结果精确、可靠。
采用RMSE分别对不同栽培模式下不同大豆节间长度、节间粗度、茎干重模型进行检验, 对其模拟值与观测值分别做1∶ 1关系图(图4、5、6)。可以看出, 不同栽培模式下不同大豆品种模拟值与实测值较接近, 在套作模式下节间长度、节间粗度、茎干重模拟的准确度R2均在0.93以上, 在单作模式下节间长度、节间粗度、茎干重模拟的准确度R2均在0.98以上。
| 表5 模型检验RMSE值 Table 5 The root mean square errors of the validated model |
 | 图4 大豆不同节间长度的预测误差Fig. 4 Prediction errors for internode length in the different varieties of soybean |
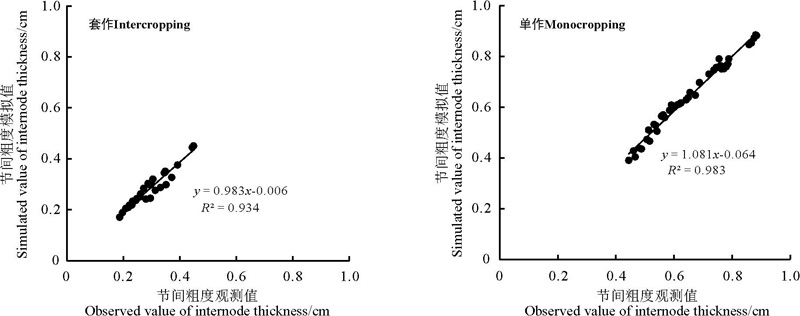 | 图5 大豆不同节间粗度的预测误差Fig. 5 Prediction errors for internode thickness in different varieties of soybean |
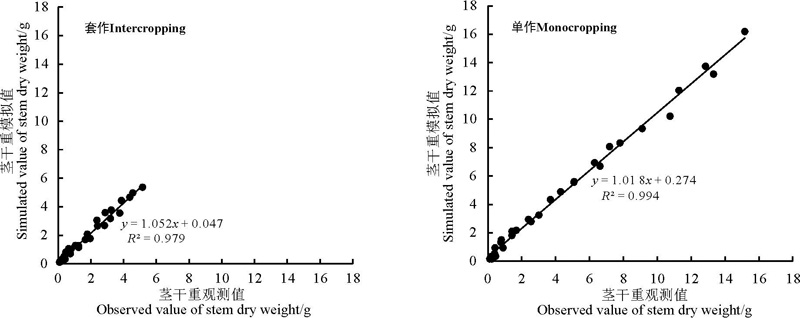 | 图6 大豆不同生理发育时期茎干重的预测误差Fig. 6 Prediction errors of the dry weight of soybean stems over the physiological growth time |
作物生长模型可以很好地描述作物生长发育的特征, 如在水稻[23]、玉米[24]、大豆[25]等作物上均有相关研究报道。但是, 对大豆生长模型的描述多侧重于单作大豆的全生育期, 而对套作大豆模型的研究鲜有报道。单作模式下, 大豆节间较短、较粗, 保持较好的形态特征; 与单作相比, 套作大豆受高秆作物玉米遮阴的影响, 光照不足, 呈避阴反应, 茎秆伸长变细; 同一品种大豆在不同栽培模式下形态有差异明显。因此本研究分别对不同栽培模式大豆茎秆生长的动态模拟, 通过量化各节间同步生长以及与生理发育时间的关系, 以固定的节间潜在长度和潜在粗度来推算模型参数, 以确定模型参数。以生理发育时间(PDT)为步长, 建立了符合大豆茎秆和生理发育时间关系的Richards方程y=α /(1+β e-γ x)1/δ , 该方程能很好地模拟大豆茎秆随生理发育时间变化的动态特征, 其方程参数有很好的生物学意义。当生理发育时间趋于无穷大时, 不同栽培模式、品种的相对最大茎长、茎粗均趋于α , α 值即为各品种的遗传参数, 由品种自身决定; β 值(初值参数)在栽培模式间变化幅度较大, 在品种间变化幅度较小; γ 值(生长速率参数)在栽培模式和品种间变化幅度均较大; δ 值(形状参数)在不同栽培模式下变异很大。通常某一大豆品种对生理发育时间的需求是一定的, 同一品种在单作和套作环境下表现出不同的形态特征。因此通过调试方程中的β 、γ 、δ 值, 对Richards方程模型进行拟合, 检验结果表明, 在单作栽培模式下, 节间长度、节间粗度观测值和模拟值的拟合较好(R2均大于0.983)。同样, 在套作栽培模式下, 节间长度、节间粗度观测值和模拟值的拟合较好(R2均大于0.934)。说明分别在不同品种及在单作、套作栽培模式下所建立的模型方程都表现出很好的拟合性, 符合茎秆生长的生物学规律, 具有一定的生物学意义。
大豆茎秆干重积累速率随着生长而变化, 呈先上升后下降趋势, 可分为渐增期、快增期和缓增期。前人的研究主要集中在大豆单作栽培模式下进行的模拟, 根据Logistic方程分析不同耕作方式对夏大豆干物质积累与分配规律的影响[26]; 根据Logistic方程建立大豆干物质累积曲线, 从而预报大豆产量[27]。然而, 在带状套作复合种植模式下对大豆茎秆干重的模拟鲜见报道。本研究结果表明, 不同的栽培模式, 3个大豆品种的全生育期的茎秆干物质积累不同, 套作大豆茎秆干重显著(P< 0.05)低于单作大豆, 由于光照不足亦导致大豆光合作用减弱, 最终使大豆茎秆干物质积累降低[9]。因此分别对单作和套作栽培模式下进行模拟, 以生长发育的实际天数为时间步长, 通过Logistic方程的拟合, 其在单作、套作栽培模式下的拟合效果较好(R2均大于0.979)。说明在单作、套作栽培模式下所建立的模型方程能很好的预测不同大豆品种生长发育实际天数与茎干重, 能较好地拟合茎秆生长动态, 符合茎秆生长的生物学规律, 具有一定的生物学意义, 将经验性和机理性有机地结合起来, 具有较好的描述性和预测性。
本研究采用单作、套作栽培模式, 3种大豆在不同栽培模式下茎秆形态建成具有较大差异的材料, 使模拟研究的可靠性更高。在系统观察的基础上, 通过对这3种不同生长习性的大豆生长进行动态模拟, 量化了各节间同步生长以及与生理发育时间的关系, 根据观测结果, 以固定的潜在节间长度和直径来推算其他模型参数。但本研究中由于仅考虑了栽培模式和品种对大豆茎秆生长的影响, 并未深入研究养分、水分等外界因子的影响, 所以需要进一步考虑温度、光照时间、养分等条件对大豆器官形态建成影响。今后将继续调试和优化模型, 考虑水肥热等因素对大豆生育进程影响, 使其在更多品种和更大地域范围内接受检验。
The authors have declared that no competing interests exist.
| [1] |
|
| [2] |
|
| [3] |
|
| [4] |
|
| [5] |
|
| [6] |
|
| [7] |
|
| [8] |
|
| [9] |
|
| [10] |
|
| [11] |
|
| [12] |
|
| [13] |
|
| [14] |
|
| [15] |
|
| [16] |
|
| [17] |
|
| [18] |
|
| [19] |
|
| [20] |
|
| [21] |
|
| [22] |
|
| [23] |
|
| [24] |
|
| [25] |
|
| [26] |
|
| [27] |
|