{kind=link}
{kind=link}
{kind=link}
{kind=link}
青藏高原垂穗披碱草种质资源遗传多样性的SSR分析
[彭语洛1  , 周青平
, 周青平2 , 陈仕勇1 , 陈有军2 , 李亚萍2 , 田莉华2 ]
, 周青平, 陈仕勇|
|


第一作者:彭语洛(1992-),女,四川宜宾人,在读硕士生,主要从事牧草种质资源研究。E-mail:[email protected]
采用SSR分子标记技术,对采自青藏高原的30份野生垂穗披碱草( Elymus nutans)种质进行遗传多样性分析。结果表明,1)筛选出的16对引物共获得了116个等位基因位点,92个多态性位点,多态性位点率(PPB)为79.75%,多态性信息(PIC)含量为0.0630.325,平均值为0.188,材料间遗传相似系数(GS)介于0.6920.976,平均值为0.828。2)聚类分析表明,在遗传相似系数0.804处,30份供试材料可聚为四大类,单独聚为一类的09-214表现出与其他材料较远的亲缘关系,主成分分析结果与UPGMA结果基本一致。群体遗传结构分析显示,大部分材料的遗传背景较为单一,群体划分的结果与聚类分析的结果部分保持一致。研究显示,供试种质材料间存在较大的差异,部分材料表现出相对独立的特性,遗传多样性较丰富,可为垂穗披碱草的保护利用、新品种的选育及优良基因的挖掘提供参考依据。
SSR markers were used to analyze the genetic diversity of 30 accessions of wild Elymus nutans collected from the Qinghai-Tibet Plateau. The results showed that: 1) A total of 116 fragments and 92 polymorphic loci were generated from 16 pairs of primers. The polymorphic loci rate (PPB) was 79.75%, with polymorphism information contents (PIC) ranging from 0.063 to 0.325 (mean 0.188). The genetic similarity (GS) coefficient ranged from 0.692 to 0.976 (mean 0.828). 2) A cluster analysis showed that at the genetic similarity coefficient of 0.804, the 30 samples clustered in four major groups. Individually clustered groups of 09-214 showed distant genetic relationships with other accessions. The result of a principal component analysis was generally consistent with those of a UPGMA. The analysis of population genetic structure indicated that most of the samples may originate from the same genetic background, and results of grouping analysis were consistent with those of the cluster analysis. Our results showed a great difference between the tested germplasm samples, with several of the accessions showing relatively independent characteristics and higher genetic diversity, which could provide reference for the protection and utilization of E. nutans, the selection of new varieties and the discovery of “good genes”.
垂穗披碱草(Elymus nutans)是小麦族(Triticeae)披碱草属(Elymus)多年生疏丛型禾草, 又名钩头草, 弯穗草, 在我国东北、华北、西北、西南等地均有野生种的分布。垂穗披碱草营养价值高、适口性好、产草量高、耐寒、耐旱、再生性和分蘖能力强[1]、耐践踏、耐瘠薄。近年来, 随着气候的变化、人类的活动及草地畜牧业的发展, 导致草原生态系统出现了不同程度的退化[2, 3]。垂穗披碱草等物种作为青藏高原的优势植物在草地恢复中发挥着重要作用[4, 5, 6, 7]。然而, 现阶段我国优质垂穗披碱草的品种数量少, 且在品质上, 无法满足当前生产所需。野生种质资源是育种的重要物质基础[8], 因此, 对垂穗披碱草野生种质资源的收集、评价是当前的重要工作之一。
迄今为止, 已有许多学者从不同的角度对垂穗披碱草种质资源及其近缘物种的遗传多样性进行了研究, 包括育成品种[9, 10], 落粒性种质资源[11], 抗旱种质资源[12], 抗寒种质资源[13], 耐盐碱种质资源[14]等, 其结果为垂穗披碱草种质资源的评价、保护和利用提供了参考依据。但我国垂穗披碱草种质资源数量大、种类繁多、且分布广泛, 随着新资源数量的增加, 垂穗披碱草遗传多样性研究需要不断的完善和补充, 以满足育种的需求。
种质遗传多样性的评价, 主要包括表型水平、分子水平等方面。相对于表型水平, 分子水平受环境因子影响较小, 重复性及稳定性较高。因此, 从分子水平对种质资源遗传多样性进行分析评价, 可靠性更高。近年来, 国内外众多学者采用SRAP标记、RAPD标记、醇溶蛋白技术等对垂穗披碱草种质资源的遗传多样性进行了研究, 陈智华等[15]用SRAP标记对采自四川、青海、甘肃、新疆的垂穗披碱草种质进行了多态性分析; 邓竹佳等[16]采用RAPD标记对来自川西北高原的垂穗披碱草进行了遗传多样性和遗传关系分析; 马啸等[17]对采自新疆、青海、四川、西藏的垂穗披碱草种质进行了醇溶蛋白多样性分析。但利用SSR分子标记研究垂穗披碱草种质资源多样性的报道较少。SSR(simple sequence repeats)分子标记既具有PFLP标记共显性遗传的优点, 又具有RAPD标记操作简单, DNA用量少等优点, 是一种进行遗传多样性研究理想的分子标记[18, 19], 目前已广泛应用于披碱草属遗传多样性的研究[20, 21, 22, 23]。为了发掘优异性状的种质资源, 补充对垂穗披碱草种质资源的研究, 本研究拟采用SSR分子标记对以青海省为主的30份垂穗披碱草种质资源遗传多样性进行研究, 旨在为垂穗披碱草野生种质资源的保护、评价及新品种选育提供参考。
30份野生垂穗披碱草种质材料采自青海、四川、西藏3省, 其中25份采自青海、3份来自四川、2份采自西藏, 编号采集地情况如表1所列。30份种质材料种植于西南民族大学青藏高原研究院牧草种质资源圃, 并对其进行了形态学鉴定。
| 表1 供试材料 Table 1 Wild E. nutans accessions analyzed in this study |
1.2.1 基因组DNA的提取及检测 随机选取每份垂穗披碱草种质材料的30粒种子, 播于试验盆中, 置于RXZ型多段可编程智能人工气候箱(宁波江南), 条件为白天22 ℃/16 h, 夜间18 ℃/8 h, 光照强度为500 mmol· (m2· s)-1, 相对湿度为70%, 生长至三叶期时, 每份材料取1015个单株新鲜健康的叶片均匀混合[24], 用液氮研磨成粉, 采用植物基因组DNA提取试剂盒(DP305)(北京天根生化)提取基因组DNA, 用1%琼脂糖凝胶电泳和超微量分光光度计(Nano DropTM Lite, Thermo)检测其浓度和纯度。
1.2.2 引物筛选及SSR分析 根据已开发的披碱草属的EST-SSR引物序列[25], 从中挑选出40对引物(由南京金斯瑞合成)。选择4份质量较高且田间表形性状差异较大的种质材料的基因组DNA对40对引物进行扩增(JY-96G型基因扩增仪)筛选, 从中选取能够扩增出清晰条带、多态性好、稳定性较高的引物用作SSR的研究。
SSR-PCR反应体系和程序。PCR反应体系:总体积20 μ L, 2× Es Tap MasterMix(Dye)10 μ L, 1 μ L上游引物, 1 μ L下游引物, 2 μ L DNA模板, 其余由ddH2O补充。PCR扩增程序为94 ℃预变性3 min, 94 ℃变性60 s, 5059 ℃退火45 s, 72 ℃延伸60 s, 共35个循环; 最后72 ℃延伸10 min, 4 ℃保存。扩增产物采用6%的变性聚丙烯酰胺凝胶电泳分离, 1× TBE电极缓冲液, 250 V电压预电泳15 min, 上样量10 μ L, 400 V电压恒压电泳1.5 h。电泳结束后, 用蒸馏水漂洗凝胶, 用染色液(0.1%AgNO3)染色14 min、显影液(0.19 g四硼酸钠+15 g NaOH+400 mL甲醛)显影直至条带清晰并用数码相机拍照保存。
1.2.3 统计与分析 根据SSR产物银染显影结果, 统计SSR扩增条带, 按在相同迁移位置上有无条带进行记录, 有扩增条带的记为1, 没有扩增条带的记为0, 由此构建0/1二元数列矩阵, 记录引物扩增条带的信息含量, 统计总扩增条带数(total number of bands, TNB)、多态性条带数(number of polymorphic bands, NPB)、多态位点百分率(percentage of polymorphic bands, PPB)及多态性信息含量指数(polymorphic information content, PIC)。利用NTSYSpc2.10[26]数据分析软件进行材料间的Dice遗传相似系数(genetic similarity coefficients, GS)的计算, 并按非加权组平均数方法(unweighted pair group method using arithmetic averages, UPGMA)进行聚类分析及主成分分析; 采用STRUCTUREV2.3.4软件进行群体遗传结构分析[27]。
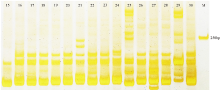
检测结果显示, 30份垂穗披碱草种质材料的DNA提取质量较高。在选择4份质量较高的垂穗披碱草种质材料的基因组DNA对40对引物进行筛选时, 共筛选出了16对条带清晰、多态性较高的SSR引物(表2)。16对SSR引物扩增结果表明:30份垂穗披碱草共检测到116个等位基因位点, 每对引物扩增出的位点变幅为515个, 每对引物扩增出的平均位点数为7.25个; 其中92个多态性位点, 引物平均的多态材料编号与表1中材料编号相同(图为15 30号)。下同。
| 表2 SSR引物序列及名称 Table 2 Sequences and names of SSR primer pairsy |
| 表3 16对SSR引物在30份供试垂穗披碱草种质材料上的PCR扩增结果 Table 3 Amplification results of the 16 SSR primers on the 30 E. nutans accessions |
性比率(PPB)为79.75%。每对引物扩增出的多态性位点变幅为311个, 每对引物扩增出的多态性平均位点数为5.75个, 引物Elw0300s019扩增出的多态性位点数最少, 为3个, 引物Elw3603s196扩增出的多态性性位点数最多, 为11个(表3), 引物Elw3264s184的扩增结果如图1所示。引物的多态性信息含量PIC值变化为0.063(Elw2202s122)0.325(Elw1420s081), 平均值为0.188。这些信息表明, 引物Elw0669s043、Elw1197s069、Elw1420s081及Elw5540s317的多态性较好、PIC值较高, 可用于日后与垂穗披碱草相关的研究。
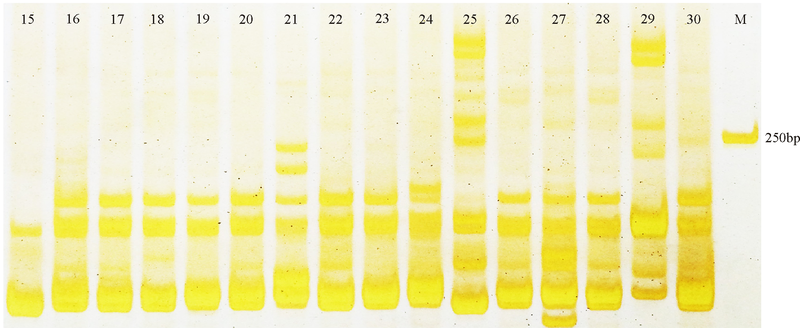 | 图1 引物Elw3264s184部分材料扩增结果Fig. 1 Amplification of the sample material using the Elw3264s184 primer |
基于16对SSR引物构建的0/1原始数据矩阵, 通过数据分析软件NTSYS-pc2.10 计算出30份供试材料间的相似系数。研究结果显示, 30份垂穗披碱草种质资源的遗传相似系数值变幅为0.6920.977, 平均值为 0.828, 其中来自海西乌兰县的I-1-1-13与来自海南共和县的I-1-1-33的相似系数最小, 为 0.692, 表明他们之间存在较大的遗传差异, 亲缘关系最远; 来自海南共和县的I-1-1-38与来自海北三角城种羊场I-1-1-39的相似系数最大, 为0.976, 表明这两份材料亲缘关系最近。这些结果表明, 供试材料之间差异明显, 遗传多样性较丰富。
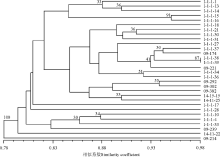
对30份垂穗披碱草种质资源进行聚类分析(图2)。聚类分析结果表明, 在遗传相似系数0.804处, 30份种质材料可聚为四大类, 其中有24份材料包含在第Ⅰ 大类中, 包括20份采自青海的材料, 2份采自西藏的材料和2份采自四川的材料。在相似系数 0.832处, 可将第Ⅰ 大类划分为 4 个亚类, 第1亚类包含5份材料, 其中2份来自果洛, 1份来自海南, 1份来自海西, 1份来自黄南; 第2亚类的组成比较混杂包含了17份材料, 4份是采自果洛, 6份是采自海南, 2份采自海北, 1份采自海西, 其中还包括采自西藏工布达江的2份材料(09-292和09-302)和2份采自四川松潘的材料(14-15-15和14-11-25); 第3亚类和第4亚类分别只包括1份材料, 分别是来自西宁湟源县的I-1-1-17和来自果洛久治县的I-1-1-28。第Ⅱ 类包括3份采自海南和1份采自海西的材料。第Ⅲ 类仅包括1份材料, 来自四川松潘的14-11-22。来自海西蒙古族藏族自治州天俊县的(09-214)单独聚为第Ⅳ 类(100%支 持率), 说明这份材料的遗传背景与其他材料存在较大的差异, 亲缘关系较远, 表现出遗传特异性, 这与其形态学的特异性相一致, 形态学表现出植株高大, 叶量丰富, 穗状花序较长等特点。
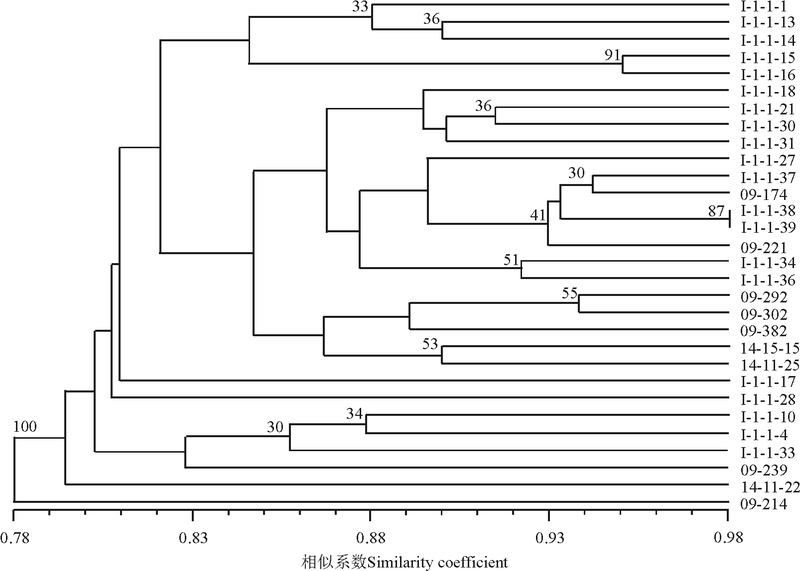 | 图2 基于SSR标记的30个供试材料的UPGMA聚类图Fig. 2 Dendrogram constructed from a UPGMA, based on SSR marker for 30 E. nutans accessions |
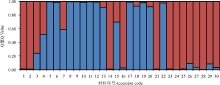
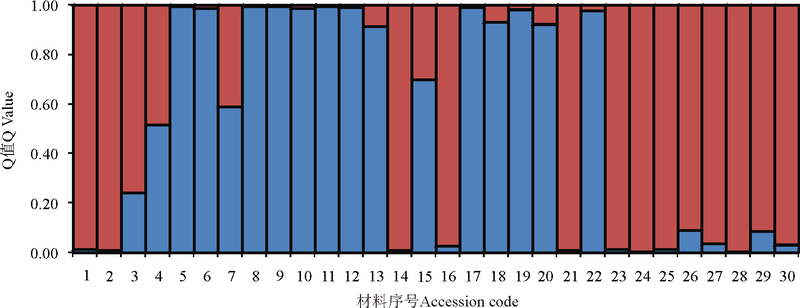
基于SSR数据对供试30份垂穗披碱草进行了群体遗传结构分析。当K=2时, △ lnP(D)具有明显的峰值, 是最适合的群体划分K值(图3)。大部分材料的遗传背景较为单一, 群体划分的结果与聚类分析的结果部分保持一致, 同时也表明本研究中的供试材料的遗传背景与其地理来源没有明显的相关性。
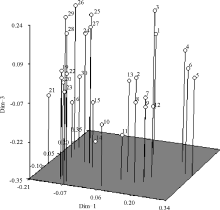
基于遗传相似系数对全部供试垂穗披碱草种质资源进行主成分分析, 其中前3个主成分的贡献率分别是82.39%、2.32%和1.90%。主成分分析三维散点图(图4)更为直观地反映其遗传亲缘关系, 位置相靠近者表示亲缘关系近, 远离者表示亲缘关系远。30份供 试垂穗披碱草种质资源主成分分析结果与聚类分析结果基本一致。
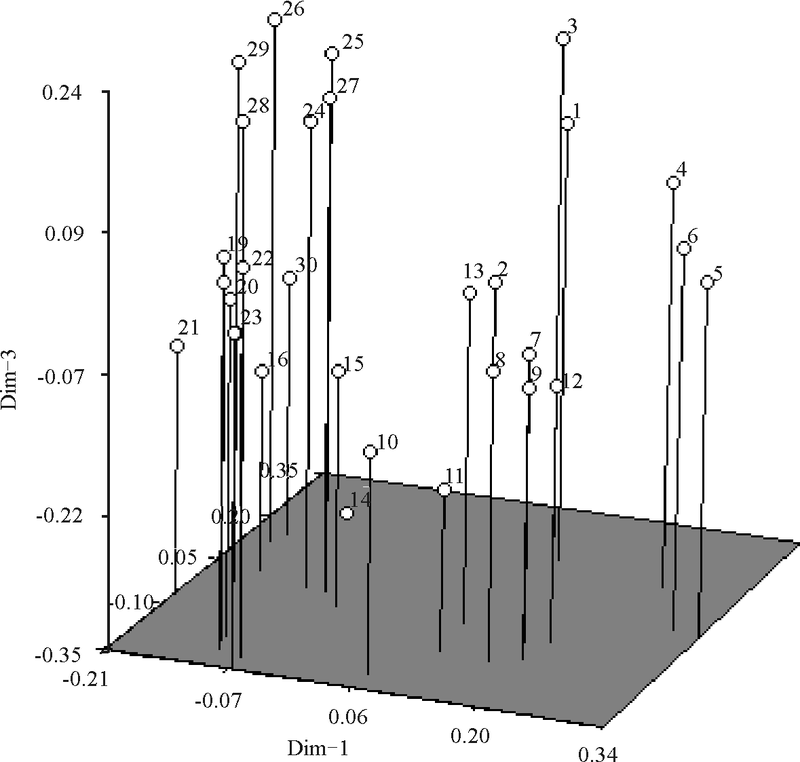 | 图4 30份垂穗披碱草种质材料依据SSR的三维主坐标分析Fig. 4 The plot of principal coordinates analysis of 30 E. nutans accession based on SSR analysis |
作为挖掘种质资源利用深度和广度强有力的手段, 分子标记已成为目前评价植物遗传多样研究的主流。目前不同类型的遗传标记已广泛应用于披碱草属种质资源的遗传评价、种间关系、遗传图谱的构建[28, 29, 30]。对垂穗披碱草遗传多样性进行研究, 可对该物种种质资源的保护利用及新品种的选育起到补充作用。本研究用SSR标记技术得到垂穗披碱草的多态性比率(PPB)为79.75%, 低于陈智华[31]用SSR标记对67份垂穗披碱草种质进行多样性分析99.4%的多态性比率, 也低于鄢家俊等[32]采用SSR标记技术对青藏高原52份老芒麦(E. sibiricus)进行遗传多样性分析86.44%的多态性比率。其原因一方面可能是由于本研究采用的是DNA单株混合提取, 从而掩盖了种质内的遗传变异, 导致决定种质间遗传关系的多态性位点数降低; 另一方面本研究供试材料样本数量有限且采集的地理范围狭窄(主要采自青海), 各材料间基因存在一定的交流。但高于张成林等[33]采用SSR标记对10份垂穗披碱草种质进行遗传多样性分析41.77%的多态性比率, 吴昊等[34] 采用SSR标记技术对35份老芒麦野生资源进行分析76.99%的多态性比率。本研究利用16对SSR引物在30份垂穗披碱草种质中共扩增出了116个等位基因, 其中92个多态性位点, 每对引物扩增出的平均位点数为7.25个, 说明了SSR分子标记对垂穗披碱草多样性研究是非常有效的, 具有多态性高的特点。多态性信息(PIC)含量为0.0630.325, 平均值为0.188, 说明了供试材料间变异程度高。
根据已报道的垂穗披碱草遗传评价结果表明, 在大空间尺度条件下, 野生的垂穗披碱草种质表现出明显的遗传和地理分化[35]。陈智华[31]、苗佳敏等[36]研究表明, 我国西部的垂穗披碱草中来自新疆和青藏高原的垂穗披碱草分化明显, 其中青藏高原地区的四川、西藏、青海等地区也存在地理分化。在本研究中, 根据聚类、群体结构及主成分等分析表明供试材料的聚类与其地理来源表现出的相关性较低, 这可能由于本研究中的材料基本上来自青海各地, 具有较为相似的气候及生境条件。这也表明垂穗披碱草在小尺度条件下的变异与其地理来源相关性不高。所以, 不同尺度的材料研究更能全面地反映该物种的遗传变异信息, 本研究为垂穗披碱草遗传多样性的研究提供了补充。
采用SSR标记对30份野生垂穗披碱草种质资源进行研究, 16对引物共获得了116个等位基因位点, 92个多态性位点, 多态性位点率(PPB)为 79.75%, 多态性信息(PIC)含量为0.0630.325, 平均值为0.188, 说明了SSR标记在垂穗披碱草多样性研究中是非常有效的, 具有多态性高的特点。种质材料间存在较大的差异, 遗传多样性较丰富, 部分材料(09-214)表现出相对独立的特性, 可为垂穗披碱草保护和利用、新品种的选育及优良基因的挖掘提供参考依据。在今后深入展开对垂穗披碱草资源的研究时, 不仅要注重样本数量的丰富度, 还要充分注重材料地理来源的多样性, 充分发掘野生种质的遗传潜力。
The authors have declared that no competing interests exist.
| [1] |
|
| [2] |
|
| [3] |
|
| [4] |
|
| [5] |
|
| [6] |
|
| [7] |
|
| [8] |
|
| [9] |
|
| [10] |
|
| [11] |
|
| [12] |
|
| [13] |
|
| [14] |
|
| [15] |
|
| [16] |
|
| [17] |
|
| [18] |
|
| [19] |
|
| [20] |
|
| [21] |
|
| [22] |
|
| [23] |
|
| [24] |
|
| [25] |
|
| [26] |
|
| [27] |
|
| [28] |
|
| [29] |
|
| [30] |
|
| [31] |
|
| [32] |
|
| [33] |
|
| [34] |
|
| [35] |
|
| [36] |
|