{kind=link}
{kind=link}
短期封育对伊犁绢蒿荒漠群落特征的影响
[李坤 , 董乙强, 安沙舟, 孔晓晶, 杨娇, 阿依敏·波拉提, 关欣]
, 董乙强, 安沙舟, 孔晓晶, 杨娇, 阿依敏·波拉提, 关欣]
, 董乙强, 安沙舟, 孔晓晶, 杨娇, 阿依敏·波拉提, 关欣]
|
|


第一作者:李坤(1993-),男,安徽萧县人,在读硕士生,主要从事草地资源与生态研究。E-mail:[email protected]
本研究以新源县和呼图壁县典型的伊犁绢蒿( Seriphidium transiliense)荒漠为研究对象,采用野外调查的方法,测定封育区和放牧区伊犁绢蒿荒漠植被生态经济类群、群落特征及其多样性,研究不同气候区荒漠群落特征对封育的响应规律,为草地资源的合理利用提供一定的基础资料和科学依据。结果表明:1)封育围栏内,草原植物群落的高度、盖度、密度和生物量有大幅度提高,显著高于围栏外( P<0.05);2)在封育围栏内,草地群落Pielou均匀度指数增大,香浓指数和Patrick指数减小,伊犁绢蒿荒漠植被恢复明显。
We studied the response of desert community characteristics to fencing in different climatic zones, which was aimed at providing certain basic information and scientific basis for the rational utilization of grassland resources. Seriphidium transiliense deserts in Xinyuan and Hutubi counties were selected as the research areas, and the eco-economic group, community characteristics, and its diversity were analyzed using a field-survey sampling method. The results showed that: 1) The height, cover, density, and biomass of the grassland community increased significantly, which was significantly higher than those outside the fence; 2) Under the fence, Pielou’s evenness index of grassland communities increased, while the fragrant index and Patrick index decreased, and the desert vegetation comprising S. transiliense recovered remarkably.
新疆是我国主要草原牧区之一, 天然草地面积辽阔, 同时也是最重要的春秋牧场。其中, 蒿类荒漠是荒漠草地的重要类型[1]。长期的过牧导致草地退化现象十分普遍, 局部地区已经极度退化或完全被破坏[2]。如何恢复退化蒿类荒漠成为亟待解决的问题。
自20世纪80年代起, 封育减少了放牧压力, 使得退化草地拥有休养生息的机会, 为植被恢复和重建提供条件[3, 4, 5, 6], 能够大幅度增加植被的盖度、平均高度、草群密度、地上生物量和草根含量等[7], 是一种有效而且简便易行的促进草原恢复的策略[8, 9]。封育是草地修复和重建的重要措施, 被世界各国广泛采用[10, 11, 12, 13]。
迄今为止, 有关研究大多集中在同一地区围栏内外草地群落构成、物种生物多样性、地上、地下生物量、不同禁牧年限、土壤理化性质、动物群落结构及多样性等对围封产生的效应[14, 15, 16], 缺少对相同草地类型在不同气候区的禁牧效果的研究, 尤其在蒿类荒漠采取的短期封育手段的研究[17]。
为此, 2017年5月起, 开展了在不同气候区(新源县和呼图壁县)春牧场围栏内外伊犁绢蒿种群特征、生态经济类群特征及其群落多样性的研究, 探讨围栏封育对该类草原的影响, 为草地资源的合理利用提供一定的基础资料和科学依据。
研究区分别位于天山北坡西段的新源县(43° 22' N, 82° 35' E)和天山中段的呼图壁县(43° 57' N, 86° 32' E), 海拔在9001 000 m。两地均属于典型伊犁绢蒿荒漠。新源县属大陆性半干旱气候, 冬暖夏凉, 降水多, 气候温凉, 年降水量400467 mm。呼图壁县属温带大陆性干旱半干旱气候, 年降水量101167 mm, 年均温6.0 ℃, ≥ 10 ℃年有效积温3 553 ℃· d。两地年均日照时数相差793.5 h, 无霜期140180 d, 土壤为荒漠灰钙土。植被均以伊犁绢蒿(Seriphidium transiliense)为建群种, 伴生种有弯果葫芦巴(Trigonella arcuata)、早熟禾(Poa annua)、猪毛菜(Salsola collina)、角果藜(Ceratocarpus arenarius)、角果毛茛(C. orthoceras)、鹤虱(Carpesium abrotanoides)、蒲公英(Taraxacum mongolicum)、黄芪(Fisch bunge)、鸢尾(Iris tectorum)、老鹳草(Geranium wilfordii)、草原苔草(Carex lioarocrpos)、莎草(Cyperus rotundus)、委陵菜(Potentilla chinensis)、木地肤(Kochiap rostrata)等。
试验采用完全随机试验设计, 分别在天山北坡西段(新源县)和中段(呼图壁县)典型的伊犁绢蒿荒漠区设置两个处理, 即封育围栏内(enclosure, EN)和自由放牧区(freely grazing, FG)。于2012年和2015年对试验区进行围栏封育, 至2017年分别封育了6和3 年, 即新源封育区封育6年, 呼图壁封育区封育3年, 均为相对短期封育, 封育面积30 000 m2。新源放牧区和呼图壁的放牧区在封育前后自由放牧区放牧强度保持不变, 且放牧率水平基本一致。
1.3.1 样地设置与野外调查 于2017年5月分别在新源和呼图壁伊犁绢蒿荒漠区进行野外调查取样。封育区和自由放牧区各设定3条样带, 每条样带间隔大于10 m, 在每条样带上设置10个1 m× 1 m的样方, 样方间距均为10 m, 共计样方120个, 测定指标包括每个样方中出现的植被密度、植物种高度、植物种类、地上生物量等。
盖度(%):采取针刺法测定。
密度(株· m-2):直接计数法记录物种株丛数。
植物种高度(cm):采用直尺测定植株的自然高度。
地上生物量(g· m-2):采用齐地面刈割法, 分种齐地刈割后装入带有标签的信封袋中带回实验室65 ℃烘至恒重后称重。
Patrick丰富度指数(P):P=S;
Shannon-Wiener指数(H):H=-∑ PilnPi;
Pielou均匀度指数(E): E=(-∑ PilnPi)/lnS;
Simpson优势度指数(D): D=1-∑
式中:S为样地总物种数量; Pi为第i个物种的重要值。
1.3.2 生态经济类群的区分 按照草地植物生态经济类群的划分方法[18], 统计伊犁绢蒿荒漠植物组成情况, 将其划分为一、二年生草本和多年生草本、半灌木、短生类短生植物。
1.3.3 数据分析 采用SPSS 20.0中的One-way ANOVA对不同处理的种群和群落数量特征进行差异性分析, 采用Excel进行数据的预处理, 运用Origin 8.0进行图形的绘制。
蒿类荒漠草地经封育后, 经济类群数量有所增加(表1)。新源县短期封育区内和自由放牧区内, 二者均出现4种菊科植物, 分别为伊犁绢蒿、鹤虱、狗娃花(Heteropappus hispidus)和蒲公英; 2种豆科植物, 分别为弯果葫芦巴、黄芪。封育区较放牧区多了1种菊科植物(蒲公英)、1种藜科植物(角果藜)和1种杂类草(莎草)。呼图壁县短期封育区内和自由放牧区相比较, 二者均出现2种豆科植物, 分别为弯果葫芦巴、黄芪; 1种禾本科植物(早熟禾); 封育区内比自由放牧区少1种菊科植物(狗娃花), 增加了3种杂类草, 分别为莎草科(草原苔草、莎草)、百合科[郁金香(Tulipa gesneriana)]; 自由放牧区增加了1种藜科植物(角果藜)。
| 表1 围栏内外植物经济类群组成 Table 1 Composition of economic groups of plants inside and outside the enclosure |
两个不同气候区, 新源出现藜科[木地肤、灰藜(Chenopodium album)], 菊科(狗娃花、蒲公英)和蔷薇科(委陵菜)。呼图壁出现藜科(猪毛菜)、十字花科[葶苈(Draba nemorosa)]、百合科(郁金香)。
优势度表示一个种在群落中的地位与作用, 植物种群优势度的大小表明植物在群落中地位的高低。封育区植物群落主要植物优势度发生明显变化, 其中苔草、莎草、鸢尾优势度明显增加, 而葫芦巴、鹤虱、早熟禾、黄芪相对下降(表2)。不管在围封区还是在放牧区, 伊犁绢蒿占据绝对优势地位, 新源和呼图壁封育区内优势度比放牧区分别增长了40.91%、24.57%。
| 表2 围栏内外物种优势度 Table 2 Dominance of species inside and outside fencing |
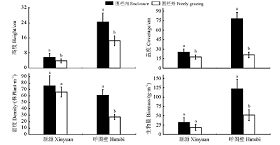
与围栏外相比, 围栏内新源的伊犁绢蒿高度、盖度、密度和生物量分别显著增加了31.79%、30.59%、12.19%和13.93%(P< 0.05)(图1), 呼图壁分别显著增加了40.59%、73.12%、55.3%、57.69%(P< 0.05), 其中呼图壁伊犁绢蒿高度、生物量增加尤为明显。在降水量低的呼图壁围栏内伊犁绢蒿高度、盖度、生物量较降水量高的新源分别增加了335.00%、215.50%和277.02%, 密度降低了19.33%。
 | 图1 封育对伊犁绢蒿种群特征的影响 不同小写字母表示同一地点封育区和自由放牧区间差异显著(P< 0.05)。下同。Fig. 1 Effect of enclosure on the diversity of Seriphidium transiliense population Different lowercase letters indicate significant difference in the same place at the 0.05 level; similarly for Table 3 and Fig. 2。 |
新源自由放牧区比短期封育区高度显著降低了24.07%(P< 0.05), 与短期封育区相比, 自由放牧区生物量降低了6.3%(P< 0.05), 但群落密度和盖度比围栏内显著增加了30.97%和22.19%(P< 0.05)(表3)。呼图壁自由放牧区比短期封育区高度降低了53.23%(P< 0.05), 生物量降低了45.62%, 盖度降低了60.62%(P< 0.05), 密度增加了19.84%(P< 0.05)。
| 表3 不同地区伊犁绢蒿荒漠草地群落特征 Table 3 Community characteristics of desert grassland of Seriphidium transiliense in different areas |
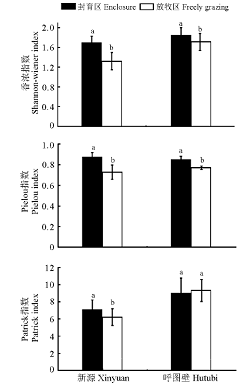
两地区短期封育和自由放牧区Shannon-Wiener指数、Pielou指数和Patrick指数的变化如图2所示。新源短期封育区比自由放牧区Shannon-Wiener指数增加了21.95%, Pielou指数增加了16.65%, Partick指数增加了12.06%(P< 0.05)。呼图壁地区封育区比自由放牧区Shannon-Wiener指数增加了7.16%, Pielou指数增加了9.23%, Partick指数降低了3.44%(P< 0.05)。新源封育区内比呼图壁, 草地群落Pielou均匀度指数增加了2.7%, Shannon-Wiener指数和Patrick指数分别减少了28.53%、30.95%。
 | 图2 围栏对伊犁绢蒿群落特征的影响Fig. 2 Effects of enclosure on the diversity of Seriphidium japonicus community |
围栏封育作为退化草地恢复治理的手段之一, 由于阻断了家畜对草原植物的采食干扰, 导致围栏内草地植物地上盖度、高度、密度和生物量显著高于围栏外, 就同一地区而言, 本研究结果也进一步验证了部分学者的报道。孙宗玖等[19]研究表明, 群落总盖度、总产量的增加主要由藜科草类(木地肤、角果藜)的增加所引起, 而密度则主要由藜科草类的角果藜及伊犁绢蒿的实生苗增加所引起。但本研究结果表明, 不同降水量的两个地区, 降水量高的地区藜科植物增加比例大, 但是群落的高度、盖度、密度和生物量均低于降水量低的地区, 这可能是荒漠植被群落中封育时间长短以及放牧强度和地表枯落物的现存量的影响所致。在自由放牧区草地由于动物采食喜好, 导致优良牧草生长受抑制, 而动物不喜好牧草受到的影响相对较小, 最终其优势度在群落中升高, 而优良牧草(伊犁绢蒿、灰藜)优势度下降。其中, 角果藜作为指示性植物出现在新源县围栏内和呼图壁围栏外。角果藜出现说明该处草地已开始退化, 这与自由放牧区的放牧压力息息相关。刘雪明和聂学敏[20]研究围栏封育措施改变退化草地群落的种类组成, 与本研究结果不尽相同。封育提高优良牧草(伊犁绢蒿、灰藜)竞争力, 影响群落生长方向, 指导群落内部间、种群间物种转变。
α 多样性是群落物种多样性、均匀度以及丰富度的测量指标, 可以反映物种间通过竞争而产生对外界环境的适应能力及群落受干扰的程度[21]。本研究表明, 随着降水量的增加, 草地群落Pielou均匀度指数增加, Shannon-Wiener指数和Pratick指数减少。赵哈林等[22]对科尔沁沙质草地植物的研究认为, 放牧强度对草地群落Shannon-Wiener指数和Pratick指数呈下降趋势, 与本研究结果一致。另一方面, 降水量的增加在一定程度上增加了物种Pielou均匀度, 这与群落物种多样性、丰富度不一致, 而具有相反趋势。主要原因是, 群落物种的多样性和丰富度程度越低, 物种对生境的分割程度就越低[23, 24], 所以导致群落物种Pielou均匀度的越高。
本研究表明, 不同气候区草原经过围栏封育后, 降水量增高不一定是主导物种丰富度和植被多样性增加的主要因素, 还可能是土壤质地以及封育效果等其他因素的影响。
The authors have declared that no competing interests exist.
| [1] |
|
| [2] |
|
| [3] |
|
| [4] |
|
| [5] |
|
| [6] |
|
| [7] |
|
| [8] |
|
| [9] |
|
| [10] |
|
| [11] |
|
| [12] |
|
| [13] |
|
| [14] |
|
| [15] |
|
| [16] |
|
| [17] |
|
| [18] |
|
| [19] |
|
| [20] |
|
| [21] |
|
| [22] |
|
| [23] |
|
| [24] |
|