{kind=link}
{kind=link}
{kind=link}
砷胁迫对黑麦草根系形态及养分吸收的影响
[李金波1  , 李诗刚
, 李诗刚2 , 宋桂龙1 , 濮阳雪华2 , 薛博晗1 , 隋永超1 ]
, 李诗刚, 宋桂龙, 濮阳雪华|
|


第一作者:李金波(1992-),男,山东济宁人,在读硕士生,主要从事植被恢复理论与技术研究。E-mail:[email protected]
黑麦草对重金属具有较强的耐性和富集能力,是一种良好的植物修复材料。为探究砷(As)胁迫下黑麦草的根系形态以及根系养分吸收特征,采用盆栽试验研究了100 mg·kg-1As胁迫下,多年生黑麦草( Lolium perenne)和一年生黑麦草( L. multiflorum)的根系形态和营养元素吸收情况,并分别对As吸收与根系形态和养分吸收做相关性分析。结果表明,多年生黑麦草总根长和总根表面积受抑制程度更加明显,分别降低47.6%和46.8%;多年生黑麦草根系As吸收能力是一年生黑麦草的1.2倍。As胁迫促进两种黑麦草根系对N、P、K、Mn元素的吸收,二者相比,多年生黑麦草根系吸收N、P、K三种大量元素受影响更大,而一年生黑麦草根系吸收微量元素Mn受影响更大。总体上看,多年生黑麦草的根系形态和养分吸收能力对As胁迫更加敏感。
Ryegrass was regarded as a plant species with potential for phytoremediation owing to its stronger heavy metal tolerance and accumulate capacities. For exploring the root morphological parameters and absorption of nutrient elements characteristic of ryegrass under arsenic stress, a pot experiment was performed. We studied the effect of 100 mg·kg-1 arsenic (As) stress on root morphological parameters and nutrient elements absorption of perennial ryegrass ( Lolium perenne) and annual ryegrass ( Lolium multiflorum), Moreover, we aimed to analyze the relationships of arsenic absorption with root morphological parameters and the content of nutrient elements. The results indicated that total length and total surface area of perennial ryegrass were reduced significantly by 47.6% and 46.8%, respectively, as compared with those of annual ryegrass. The absorbing capacity of As of perennial ryegrass root was 1.2 times greater than that of annual ryegrass. As stress promoted the absorption of N/P/K/Mn in two ryegrass species. Perennial ryegrass root had a greater effect on N/P/K absorption, whereas annual ryegrass root had a greater effect on Mn absorption. The root morphological parameters and nutrient absorption of perennial ryegrass was more sensitive to arsenic stress.
砷(Arsenic, As)是生物生长的非必需元素, 也是国际癌症研究组织确认最早的一级致癌物, 其在环境中的平均浓度达到5 mg· kg-1 [1]。目前砷污染已经成为一个非常重要的环境问题, 在我国湖南、河南等省份均有不同程度的砷污染问题, 并曾出现砷污染地下水导致人中毒事件的发生[2]。植物在砷污染的土壤中生长, 生长发育会受到抑制, 生物性状和养分吸收特征也会发生异常变化[3]。
治理土壤重金属污染的措施主要有工程治理技术、农业改良措施、生物修复技术、植物-化学修复技术[4]。生物修复技术中的植物修复技术具有成本低、对环境扰动小等优点, 具有极大的应用前景[5]。植物修复技术的关键是寻找好的植物修复材料, 黑麦草(Lolium perenne)具有生物量大、生长快、易于种植等特点, 且有报道称黑麦草对重金属有较强的抗性和富集能力[6], 是一种良好的植物修复材料。万玉山等[7]在黑麦草、芥菜(Brassica juncea)、苜蓿(Medicago sativa)和高羊茅(Festuca arundinacea)4种植物之间进行了Cd、B[α ]P污染土壤修复植物的筛选, 结果表明黑麦草和芥菜对土壤中有效态Cd均有富集能力, 黑麦草对污染土壤的修复效果强于苜蓿和高羊茅。李松克等[8]研究了多年生黑麦草对黄壤重金属污染土壤的修复, 结果表明多年生黑麦草对Cd、Cr、Pb、As、Hg都有一定的富集能力。
植物在受到重金属胁迫后, 其根、茎、叶都会吸收土壤中的重金属。但由于根系直接接触土壤或水环境, 所以根系是植物吸收重金属的主要器官[9]。同时, 根系也是植物吸收养分和水分的主要器官, 植物地上部的生长发育会受到根系形态直接或间接的影响[10]。当植物受到重金属Cd胁迫时, 各器官中的Cd含量一般为根、叶、茎、花、果、籽粒, 所以Cd对植物的毒害首先表现在根系形态和生理功能的改变[11, 12]。陈伟等[13]研究了重金属离子对两种草坪草荧光特性和根系形态的影响, 结果表明高羊茅总根长、总根表面积、总分叉数、总根尖数都显著低于未经过重金属处理的对照, 多年生黑麦草则表现为对低浓度金属离子抗性较强, 高浓度金属离子抗性较差且二者都有随浓度升高而降低的趋势。朱云集等[14]研究了As胁迫对小麦(Triticum aestivum)根系生长及活性氧代谢的影响, 也得出了类似的结果。重金属对植物的危害之一就是影响营养元素的吸收, 使植物体内养分离子失衡[15]。As对冬小麦根系生长和养分吸收的影响研究结果表明, 当砷浓度为5~10 mg· L-1时, 小麦根系中P、K、Mg、Mn、Cu和Zn的含量有增加的趋势, 当砷浓度为15~20 mg· L-1时, 小麦根系中P、Mn、Zn的含量则相比对照有降低趋势[16]。综上所述, 虽然已有一些重金属对植物根系形态和养分吸收影响的研究, 但在As胁迫下对黑麦草的根系形态和养分吸收的研究却鲜见报道。因此, 本研究以一年生黑麦草和多年生黑麦草为材料, 研究其在As胁迫下的根系形态和根系养分吸收特征, 以探究As胁迫对黑麦草根系形态和养分吸收的影响, 并试图分析两种黑麦草之间的差异。
多年生黑麦草(Lolium perenne)和一年生黑麦草(L. multiflorum)种子均来自北京正道种业有限公司, 品种分别为Mathilde和Idyll。
供试土壤取自北京林业大学草坪研究所昌平试验基地(116.25° E, 40.18° W), 土壤类型为潮土, 质地中壤, pH为7.68, 全氮含量1.33 g· kg-1, 速效钾含量70.2 mg· kg-1 , 有效磷含量13.9 mg· kg-1 , 有机质含量8.2 g· kg-1, As含量0.337 mg· kg-1 。将土壤风干并过5 mm孔径筛, 按照14∶ 1(w∶ w)混入腐熟鸡粪作为底肥。
盆栽试验在北京林业大学草坪研究所昌平试验站进行, 试验选用上内径20 cm、下内径10 cm、高28 cm的锥形塑料花盆, 每盆装混合土3 kg。As以NaAsO2配成200 mL溶液浇入土壤, 静置两周。
试验采取完全随机区组设计, 共设0(CK)、100 mg· kg-1 两个As浓度(以纯As∶ 混合土计), 分别用DCK、D100、YCK、Y100表示多年生黑麦草的CK和As处理, 一年生黑麦草的CK和As处理。各处理设置3次重复, 共计12盆。
在育苗盘里育苗, 待出苗10 d后选取长势一致的幼苗移栽至As处理花盆, 生长适应2周后定株10株, 然后定期定量培养60 d。
参考Green等[17]的方法, 将植物根系取出并清洗干净, 将根系剪下后随机选择一株平铺在根系扫描盘内, 采用Epson Scan V700根系扫描仪扫描根系图像, 将图像保存至计算机并使用WinRHIZO PRO 2013根系分析系统软件(Regent Instruments Inc., Canada)计算总根长、总根表面积、根平均直径、总根体积、根尖数、分枝数。
根系扫描后全部收集置于80 ℃下烘干至恒重, 烘干样品使用粉碎机粉碎并过0.3 mm孔径筛备用。首先称取0.200 g, 使用Hanon220s石墨消解仪采用H2SO4-H2O2消化完全后定容至100 mL, 然后用全自动流动分析仪(SEAL AA3)测定全N含量。再称取0.500 g, 采用HNO3-H2O2消化完全后动容至50 mL, 采用ICP-MS(安捷伦 7700)测定As、P、K、Ca、Mg、Mn含量。
原始数据经Excel 2010软件整理, 运用SPSS for Windows 17.0软件对黑麦草根系形态参数、As及养分含量进行单因素方差分析并将As含量分别与根系形态及养分含量做相关性分析, 分析结果用origin作图。
As胁迫下多年生黑麦草和一年生黑麦草的根系形态变化如图1所示。当As浓度为100 mg· kg-1时, 两种黑麦草的总根长都受到抑制, 但一年生黑麦草并未达到显著水平(P> 0.05), 二者分别比各自对照降低47.6%和17.7%。一年生黑麦草的总根表面积在As胁迫下几乎无差异, 而多年生黑麦草则显著降低了46.8%(P< 0.05)。一年生黑麦草的根平均直径和总根体积在As胁迫下分别比对照增加23.8%和24.3%, 多年生黑麦草的总根体积则比对照显著降低46.1%(P< 0.05)。两种黑麦草的根尖数和分枝数在As胁迫下均显著降低(P< 0.05), 根尖数分别降低56.8%和55.0%, 分枝数分别降低67.1%和44.1%。总体来看, As胁迫下一年生黑麦草的根系受抑制程度要明显小于多年生黑麦草, 这也说明一年生黑麦草根系的As耐性要优于多年生黑麦草。
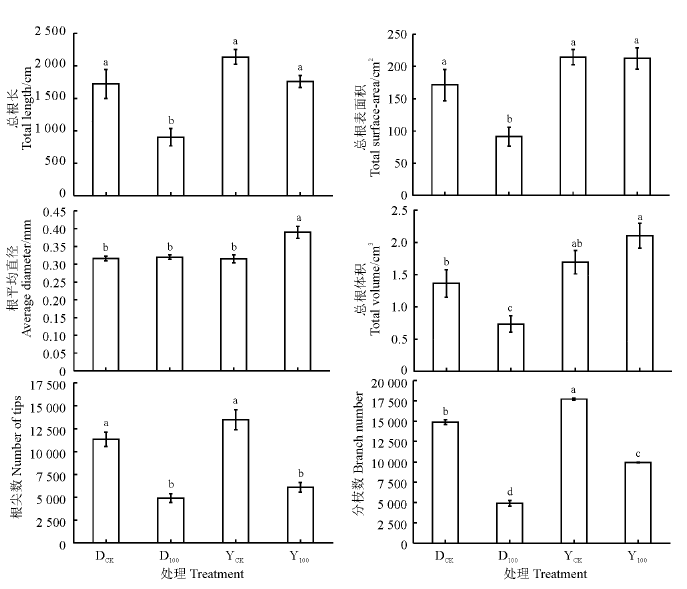 | 图1 As胁迫下两种黑麦草根系形态变化 不同小写字母表示处理间差异显著性(P< 0.05), DCK、YCK分别表示多年生黑麦草、一年生黑麦草的对照, D100、Y100分别表示多年生黑麦草、一年生黑麦草100 mg· kg-1砷处理, 下同。Fig. 1 Changes in root morphological features of two ryegrass species under arsenic (As) stress Different lowercase letters indicate significant difference between the four treatments at the 0.05 level; DCK, YCK indicate the blank for Lolium perenne and L. multiflorum respectively, D100、Y100 indicate the 100 mg· kg-1 As treatment for L. perenne and L. multiflorum respectively; similarly for the following figures. |
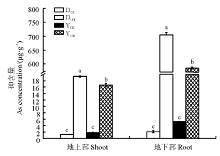
两种黑麦草在As胁迫下地上部和地下部As含量都显著升高(P< 0.05)(图2), 多年生黑麦草地上部As含量是一年生黑麦草的1.16倍(P< 0.05), 二者分别为各自对照的15.36和9.15倍; 多年生黑麦草地下部As含量是一年生黑麦草的1.20倍(P< 0.05), 二者分别为各自对照的337.89和112.62倍。多年生黑麦草对As的吸收能力明显优于一年生黑麦草, 这也可能是多年生黑麦草根系受抑制程度更大的原因。
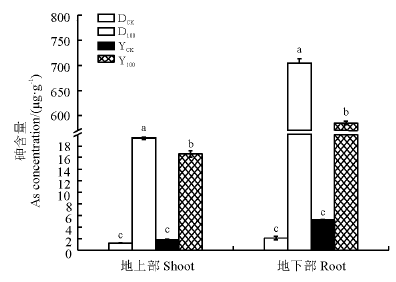 | 图2 As胁迫下两种黑麦草As含量Fig 2 As concentration in two ryegrass species under As stress |
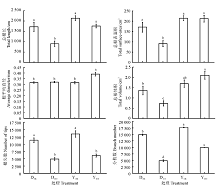
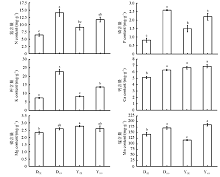
两种黑麦草根系对营养元素的吸收都受到了As胁迫的影响(图3)。两种黑麦草在As处理下N、P、K都呈现增高趋势, 并且除一年生黑麦草N含量之外都达到显著水平(P< 0.05), As处理下二者N含量分别比对照增高了125.7%和29.4%, P含量分别比对照增高了216.9%和47.6%, K含量分别比对照增高了207.0%和65.6%。多年生黑麦草Ca含量较对照显著增高了22.6%(P< 0.05), 一年生黑麦草几乎无差异(P> 0.05)。两种黑麦草在As处理下Mg含量都没有显著差异(P> 0.05)。而Mn含量都显著增高(P< 0.05), 二者分别比对照增高了19.8%和58.2%。总体来看, As处理促进了两种黑麦草根系对养分元素的吸收, 但两种黑麦草之间存在较大差异。
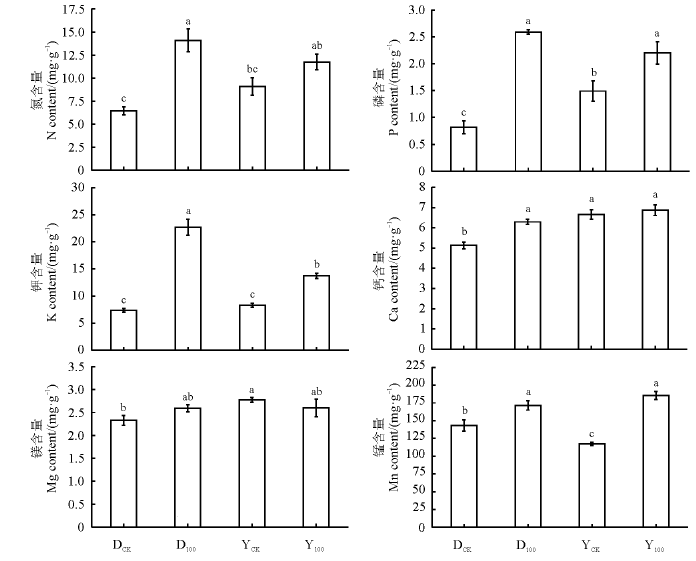 | 图3 As胁迫下两种黑麦草根系N、P、K、Ca、Mg、Mn含量Fig. 3 N, P, K, Ca, Mg, and Mn concentration in the roots of two ryegrass species under As stress |
在没有As处理时, 一年生黑麦草根系对于N、P、K、Ca、Mg五种大量元素的吸收大于多年生黑麦草, 除N和K之外都达到了显著水平(P< 0.05); 一年生黑麦草对于Mn元素的吸收要小于多年生黑麦草并且二者差异显著(P< 0.05)。结果说明As胁迫对于根系吸收N、P、K的影响多年生黑麦草大于一年生黑麦草, 而对于根系吸收Mn的影响一年生黑麦草大于多年生黑麦草。
As胁迫下, 两种黑麦草在As与6种营养元素吸收的相关性方面, 表现出了趋势基本一致, 但显著性明显不同(表1)。对于多年生黑麦草而言, As与N、P、K、Ca四种元素极显著正相关(P< 0.01), 与Mn元素呈现显著正相关关系(P< 0.05), As与Mg元素没有显著相关性。对于一年生黑麦草而言, As与K、Mn两种元素极显著正相关(P< 0.01), 与N、P、Ca、Mg四种元素没有显著相关性。总体来看, As与6种营养元素基本呈现为正相关关系, As处理促进了两种黑麦草根系对于营养元素的吸收, 但二者之间表现出较大差异, 同样的As处理条件下多年生黑麦草根系养分吸收受影响更大。
| 表1 两种黑麦草根系营养元素含量与砷含量的相关系数 Table 1 Correlation coefficients between nutrient elements and As in the root of two ryegrass species |
对于多年生黑麦草而言, As含量与总根长、总根表面积、总根体积、根尖数、分枝数极显著负相关(P< 0.01)(表2)。对于一年生黑麦草而言, As与根尖数和分枝数显著负相关(P< 0.05)。两种黑麦草根系As与根平均直径都没有显著相关性, 且一年生黑麦草根系As与总根长、总根表面积、总根体积也没有显著相关性。总体来看, As胁迫抑制了两种黑麦草根系的生长, 但多年生黑麦草根系受抑制程度更加严重。
| 表2 两种黑麦草根系形态与砷的相关系数 Table 2 Correlation coefficients between root morphological parameters and As concentration in the roots of two ryegrass species |
黑麦草是常见的禾本科植物, 具有生物量大、根系发达的特点。根系是植物最早接触重金属的器官, 也是阻拦重金属的第一道屏障。有研究证明黑麦草能够有效富集土壤中的Cd离子, 并且Cd主要集中在根系中[18]。研究中两种黑麦草根系中都有较高的As含量, 这与前人的研究结果也基本一致。两种黑麦草相比, 多年生黑麦草根系As含量大于一年生黑麦草, 说明多年生黑麦草对As的吸收能力大于一年生黑麦草。植物在吸收重金属后会将大量重金属富集在根部, 较少部分转运到茎叶部分, 其原因可能是植物自身防止重金属对叶片光合作用的影响以及对茎叶新陈代谢的毒害, 这也是植物对逆境的一种适应机制[19], 相关研究证明Cd和Pb会分别降低东南景天(Sedum alfredii)[20]的叶绿素含量和破坏绢毛委陵菜(Potentilla sericea)[21]的叶绿体膜系统, 说明重金属转运到叶片后会影响植物的光合作用。
根系是植物重要的营养器官, 能够从土壤中吸收水分及养分。当土壤受到污染时, 根系也是最早感知土壤非生物逆境胁迫的器官, 根系形态能够直观反映根的适应能力[22]。结果表明, As胁迫抑制两种黑麦草根系的总根长、总根表面积、根尖数、分枝数, 对根平均直径有一定促进作用, 这与前人在小麦(Triticum aestivum)和杞柳(Salix integra)上的研究结果相一致[23, 24]。两种黑麦草之间存在较大差异, 这说明黑麦草根系对As的耐性与其基因型有关。总根长、总根表面积和总根体积能反映植物根系的吸收能力, 根尖数和分枝数则反映植物侧根的发生情况。As可能通过破坏根细胞结构、抑制根细胞分裂导致总根长、总根表面积、总根体积的减少。也有研究表明, 紫花苜蓿(Medicago sativa)根系在10 mg· kg-1 Cd胁迫下根平均直径相对CK会显著降低[22], 与本研究结果相反, 这可能与金属离子种类以及植物种有关。研究中两种黑麦草在100 mg· kg-1 As胁迫下根尖数和分枝数度显著降低, 与田晓锋等[25]在高浓度Cd下对梧桐(Firmiana platanifolia)的研究结果相类似, 但在低浓度Cd下根毛会增多。
重金属对植物的危害之一是影响植物对营养元素的吸收, 扰乱其体内的养分平衡, 进而对新陈代谢产生影响。结果表明, 两种黑麦草根系As含量与营养元素含量总体呈现出正相关关系, 二者表现出协同作用。两种黑麦草之间存在较大差异, 多年生黑麦草根系As含量与N、P、K、Ca、Mn五种营养元素含量都呈现出显著或极显著正相关关系, 而一年生黑麦草根系As含量只与K、Mn含量呈现极显著正相关关系, 说明植物因基因型不同, 重金属含量与营养元素之间的相关性也存在差异, 这与前人在6种水草上的研究结果相类似[26]。研究中2种黑麦草根系在100 mg· kg-1 As胁迫下会增加对营养元素的吸收, 其原因可能是100 mg· kg-1的As胁迫导致两种黑麦草根系生物量降低, 但是没有影响到根系对营养元素的吸收能力, 所以造成根系中养分的“ 浓缩效应” [16], 类似结果在菜豆(Phaseolus vulgaris)的研究中也有体现[27]。但是刘全吉等[16]的研究结果表明冬小麦在5~10 mg· L-1 As处理下根系中K、Mg、P、Mn、Cu含量有不同程度的提高, 在15~20 mg· L-1 As处理下根系中Mn、Cu、Zn、Fe等微量元素的含量显著降低, 这说明植物根系中As含量与营养元素的相关性不仅与植物种有关, 也与As处理浓度有关, 因此有必要在多浓度梯度As胁迫下研究两种黑麦草根系As吸收与营养元素的相关性。另外, 重金属与营养元素的关系错综复杂, 有待在分子水平上作进一步研究。
综上所述, As胁迫能抑制两种黑麦草根系的总根长、总根表面积、根尖数、分枝数, 但二者对As的耐性不同, 一年生黑麦草根系耐性优于多年生黑麦草, 多年生黑麦草总根长和总根表面积受抑制程度更加明显; 两种黑麦草对As的吸收能力存在显著差异, 多年生黑麦草优于一年生黑麦草。As促进了两种黑麦草根系对N、P、K、Mn元素的吸收, 多年生黑麦草根系中As对N、P、K 3种大量元素的促进作用大于一年生黑麦草, 一年生黑麦草根系中As对微量元素Mn的促进作用大于多年生黑麦草。两种黑麦草相比, 多年生黑麦草对As胁迫更加敏感, 其根系形态和养分吸收受到更大影响。
The authors have declared that no competing interests exist.
| [1] |
|
| [2] |
|
| [3] |
|
| [4] |
|
| [5] |
|
| [6] |
|
| [7] |
|
| [8] |
|
| [9] |
|
| [10] |
|
| [11] |
|
| [12] |
|
| [13] |
|
| [14] |
|
| [15] |
|
| [16] |
|
| [17] |
|
| [18] |
|
| [19] |
|
| [20] |
|
| [21] |
|
| [22] |
|
| [23] |
|
| [24] |
|
| [25] |
|
| [26] |
|
| [27] |
|