


{kind=link}
{kind=link}
{kind=link}
青海省东部地区熊蜂物种多样性分析
[王磊1, 2, 3  , 赵芳
, 赵芳4 , 陈振华4 , 杨俊东1, 2, 3 , 苏建平1, 2 , 张同作1, 2 , 林恭华1, 4 ]
, 赵芳, 林恭华]
|
|


第一作者:王磊(1991-),男,陕西商洛人,在读硕士生,主要从事熊蜂生态学研究。E-mail:wanglei02017@outlook.com
采用形态学和基于线粒体16S rDNA基因的分子生物学方法,对采自青海省东部地区20个县(市)的熊蜂( Bombus spp.)进行了物种鉴定及组成分析,并探讨了物种多样性与环境因子之间的关系。结果表明,在20个采样点共采集到3 195份熊蜂标本,鉴定为23个物种,隶属10个亚属;所有熊蜂的总体Shannon-Wiener多样性指数为2.67,总体Pielou均匀度指数为0.85,具有较高的物种多样性水平;有5个物种[兰州熊蜂( B. lantschouensis)、密林熊蜂( B. patagiatus)、明亮熊蜂( B. lucorum)、火红熊蜂( B. pyrosoma)和黑尾熊蜂( B. melanurus)]具有商业开发的潜力,其数量占总标本数的17.9%;Pearson相关分析和多元线性回归分析表明,年均温是影响熊蜂物种多样性的关键因素;随着温度的降低,物种丰富度、Shannon-Wiener多样性指数和Pielou均匀度指数都显著下降。本研究结果为了解、利用和保护青藏高原熊蜂生物多样性提供了关键基础资料。
, Zhao Fang, Lin Gong-hua
Using morphological characteristics and mitochondrial 16S rDNA barcoding, we identified bumblebee ( Bombus spp.) samples collected from 20 counties of eastern Qinghai Province, Qinghai-Tibet Plateau (QTP). We also investigated the species composition of bumblebees in each sampling site and analyzed the relationship between species diversity and related environmental factors. A total of 3 195 specimens was collected from 20 sampling sites, of which 23 species belonging to 10 subgenera were identified. The total Shannon-Wiener diversity was 2.67 and Pielou evenness index was 0.85, showing a high level of species diversity. Five species ( B. lantschouensis, B. patagiatus, B. lucorum, B. pyrosoma, and B. melanurus) possibly have the potential for economic development, comprising 17.9% of the total sample. The results of Pearson correlation and multiple linear regression analysis showed that the annual temperature was the key factor influencing species diversity, and as the temperature decreased, species richness, Shannon-Wiener diversity, and Pielou evenness all significantly decreased ( P<0.05). Our research provides fundamental information for the understanding, utilization, and conservation of bumblebee diversity in the QTP.
熊蜂(Bombus spp.)是膜翅目(Hymenoptera)蜜蜂科(Apidae)熊蜂族(Bombini)下的一个单系类群, 全世界约250种, 隶属于15个亚属[1]。作为野生植物和农作物的重要传粉昆虫, 熊蜂在维护生态系统平衡和提高农业效益中发挥着十分重要的作用[2, 3]。与其它传粉昆虫(包括家养的蜜蜂)相比, 熊蜂具有传粉效率高、耐阴暗潮湿环境、不易撞棚和飞逃等优点, 在设施农业中具有明显优势。目前, 在欧洲各国、美国、日本等发达国家, 熊蜂授粉技术已形成完整的产业链, 取得很大的经济效益[3]。得益于熊蜂在生态和农业方面的巨大价值, 国外的科研人员对熊蜂的分类、生理、遗传、行为和保护等学科领域, 开展了大量的研究工作[4]。
中国是熊蜂资源最丰富的国家, 约占全世界熊蜂资源的50%[5]。20世纪70年代以来, 国内外研究人员对中国熊蜂资源开展了相对系统的调查研究, 取得了比较丰硕的成果。例如, An等[6]在前人的研究基础上进行梳理, 确认中国北方地区共有77种熊蜂:北京17种、天津5种、河北25种、山西28种、内蒙古28种、陕西30种、甘肃56种和宁夏29种。Williams等[7]对中国川渝地区开展熊蜂调查和馆藏整理发现, 这一地区共有56种熊蜂。此外, 研究人员对西藏(42种)、云南(27种)、吉林(12种)和浙江(10种)等地区的熊蜂资源也有不同程度的研究[8, 9, 10, 11]。
青藏高原是全世界熊蜂生物多样性最丰富的地区之一[6]。由于生态环境恶劣, 这一地区传粉昆虫种类较少, 尤其在高寒草地生态系统中, 许多植物都严重依赖熊蜂授粉[12]。当前, 国内外科研人员先后对西藏、四川、甘肃等省(区)熊蜂物种多样性进行了调查[5, 6, 7, 8, 13]。然而, 对于青藏高原的主体省份— — 青海省, 目前没有熊蜂方面的系统性研究。青海省东部地区, 是青藏高原重要的农牧交错地带, 地形复杂、气候多变。为此, 本研究针对这一地区的熊蜂资源开展调查, 并进行物种多样性分析, 以期为了解青藏高原的生物多样性特征提供关键基础资料。
研究区涵盖青海省东部5个地级市:西宁市、海东市、海北州、海南州、黄南州, 共辖23个县市。本研究除了海东市下辖的3个县(平安、循化、民和)未找到合适的采样点外, 其他县(市)均完成一个种群的采样(表1)。
| 表1 熊蜂采样点信息和样本量 Table 1 Geographical distribution and sample size of bumblebees |
2017年6-9月, 在青海省东部地区进行采样调查, 采集了样点方圆1 km范围内的样品。采样工具为直径40 cm的捕虫网, 将捕捉到的熊蜂置于扎孔的矿泉水瓶中, 待其自然死亡, 记录详细的采集地点(经度、纬度和海拔)、采集时间和栖息地类型等信息。未死亡个体则带回实验室, 置于烘箱56 ℃条件下20 min左右, 促其死亡。熊蜂死亡后, 用昆虫针固定在昆虫盒里并逐一登记。
对所采集的标本首先按照形态特征进行归类, 依据为熊蜂胸部和腹部体节的毛的颜色及其配置、毛的长度和浓密程度, 同时参考体型大小、喙的长度、单眼数量、胸部背面和腿胫节有无绒毛覆盖等特征。将归类的熊蜂与文献中的图谱资料[5, 6, 7, 14]进行比对, 初步确定种类。对于分子鉴定, 每个形态鉴定的种类随机选取1~3头, 对其线粒体16S rDNA基因进行PCR扩增, 扩增产物进行Sanger测序, 实验条件参考已发表的文献[15]。测序结果用Chromas软件批量输出, 并结合使用MEGA软件[16]进行比对和校对。将校对后的序列在GenBank中进行在线BLAST, 下载与本研究所测序列亲缘关系最近的参考序列, 在MEGA软件中统计序列相似度。对于所测序列与参考序列完全相同的标本, 经形态比对无明显差异的, 直接判定为参考序列对应的物种。而与参考序列不完全相同的标本, 则根据文献记录[6, 7], 下载形态上相似物种的16S rDNA序列进行比对, 统计差异位点数, 利用排除法鉴别物种。
用3个统计量来描述物种组成特征:物种丰富度(Richness=S)、Shannon-Wiener多样性指数(H'=-
 | 图1 有开发潜力的5种熊蜂工蜂标本Fig. 1 Workers of five economical bumblebee species |
将鉴定后的数据输入Excel表中, 导入SPSS软件, 统计不同种群和不同物种的数量组成信息。利用R语言的vegan和spaa软件包, 计算各种群及总体的物种丰富度、Shannon-Wiener多样性指数和Pielou均匀度指数。为了探讨物种多样性与环境因子之间的关系, 用ArcGIS软件提取了各采样点年均温(annual temperature, AT)、年降水(annual precipitation, AP)、年净初级生产力(annual net primary production of the vegetation, ANPP)等信息。其中, 年均温和年降水的底图来自于青藏高原科学数据中心(http:∥www.tpedatabase.cn), 为1961-1990年国内各台站数据经插值计算得到。而年净初级生产力底图来自蒙大拿大学(ftp:∥ftp.ntsg.umt.edu), 为MODIS遥感数据2000-2015年的均值数据经模型换算而来, 计量单位为每年每平米固碳的克数。用SPSS软件将3个多样性指数与3个地理参数(经度、纬度、海拔)和3个环境因子(年均温、年降水、年净初级生产力)进行Pearson相关性分析。需要指出的是, 为了排除这些自变量之间可能存在的自相关, 用多元线性回归法的Backward策略进行进一步分析, 确定与因变量相关性最大的自变量。
青海东部地区的20个县市, 共采集到3 195头熊蜂(表2)。所有标本进行形态和分子鉴定, 结果表明, 青海省东部地区有熊蜂23种, 涵盖10个亚属。由于每个物种的1~3条序列完全相同, 每个物种仅选取1条序列进行分析。经比对和校对, 得到23条线粒体16S rDNA序列, 长度范围525~536 bp。所测序列已提交到GenBank, SP01~SP23对应GenBank号:MG681141~MG681163。由于黑尾熊蜂、火红熊蜂、兰州熊蜂和伪猛熊蜂的参考序列较短, 因此仅对比对上的区段进行分析(表2)。其中, 20个物种的序列与参考序列完全相同, 可以直接确定物种; 而有3种与斯熊蜂、锈红熊蜂和亚西伯熊蜂的参考序列分别有4、3和7个碱基(bp)的单核苷酸变异(SNP)或插入缺失(INDEL), 经过与形态上相似物种进行序列比较, 用排除法完成鉴定。亚西伯熊蜂、红束熊蜂和克什米尔熊蜂数量最多, 分别占总数的12.4%、11.9%和10.6%, 分布也较为广泛, 在绝大多数县市(≥ 16)都有分布。越熊蜂、牛拟熊蜂和锈红熊蜂则数量最少(表2、图2)。
| 表2 熊蜂物种鉴定结果及数量分布 Table 2 Species identification results and distributions of bumblebee |
 | 图2 熊蜂各物种的频率分布Fig. 2 Frequency of the bumblebee speciesty |
20个样点中, 物种数最多的是尖扎和乐都, 分别采集到熊蜂20种和16种。物种数最少的是湟中和祁连, 分别只有3个和4个物种(表3)。所有样品的总体Shannon-Wiener多样性指数为2.67, 有4个样点(乐都、贵德、尖扎、同仁)的Shannon-Wiener多样性指数大于2.0, 有2个(泽库、湟中)则小于1.0。所有样品的总体Pielou均匀度指数为0.85, 有5个样点(同仁、贵德、乐都、化隆、大通)的均匀度指数大于0.80, 而有2个(贵南、泽库)均匀度指数小于0.60。
| 表3 熊蜂物种多样性分析 Table 3 Diversity of bumblebees |
Pearson相关分析显示, 物种丰富度与经度显著相关, Shannon-Wiener多样性指数与年均温显著相关, Pielou均匀度指数与海拔显著负相关, 其余自变量和因变量之间无显著关系(P> 0.05)。西宁采样点的3个多样性指数值都异常偏离线性回归曲线, 将此采样点数据去除后重新进行Pearson相关分析; 结果显示, 物种丰富度与经度和气温同时显著相关, Shannon-Wiener多样性指数和Pielou均匀度指数都分别与海拔和气温同时显著相关, 而其余自变量和因变量之间无显著关系(P> 0.05)(表4)。
| 表4 熊蜂物种多样性指数与环境因子的Pearson相关分析 Table 4 Pearson correlation between bumblebee species diversity and environmental factors |
多元线性回归显示, 包括西宁采样点时(n=20), 最优模型中经度、年均温、海拔分别作为最优自变量与物种丰富度、Shannon-Wiener多样性指数、Pielou均匀度指数相关。而去除西宁采样点后(n=19), 年均温都作为最优自变量与上述3个因变量显著相关(表5、图3)。
| 表5 青海东部地区熊蜂物种多样性与环境因子的多元线性回归分析的最适模型 Table 5 Best fit models of multiple linear regression between bumblebee species diversity and environmental factors in eastern Qinghai |
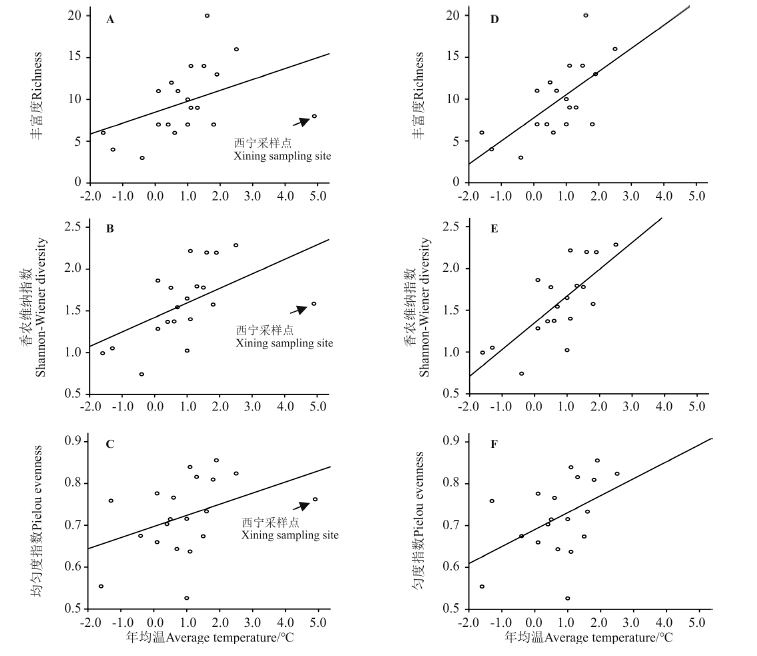 | 图3 年均温与熊蜂物种多样性指数的线性回归 A-C, 包含西宁采样点, 即n=20; D-F, 排除西宁采样点, 即n=19。Fig. 3 Linear regression between annual temperature and bumblebee species diversity A-C, including Xining sampling site, i.e. n=20; D-F, excluding Xining sampling site, i.e. n=19. |
本研究结合形态特征和分子序列分析对熊蜂进行了物种鉴定。分子序列分析的结果显示, 有20个物种的序列与GenBank中的参考序列完全一致, 结合形态特征, 可以鉴定到种。另外3个疑似物种(亚西伯熊蜂、锈红熊蜂和斯熊蜂)与参考序列有少量差异。根据此前的文献报道[6, 7], 这3个物种的毛色及其配置方面具有比较明显的鉴定特征:亚西伯熊蜂胸部前后都为黄色, 中部及翅根周围为完整的黑色条带, 腹部第3背板黑色, 尾部红色; 锈红熊蜂的胸部、翅根周围、腹部背面都是橙色; 斯熊蜂腹部第3背板黑色, 而第5背板浅橙色。亚西伯熊蜂与昆仑熊蜂(B. keriensis)在毛色上相似度较高[6], 本研究所测序列与亚西伯熊蜂的参考序列有7 bp差异(5个SNP和2个INDEL), 然而与昆仑熊蜂参考序列(No. DQ788041)之间却有48 bp差异(482/530)。锈红熊蜂与牧场熊蜂(B. pascuorum)、斯氏熊蜂(B. schrencki)、角熊蜂(B. tricornis)以及黑足熊蜂(B. atripes)在棕红色毛上有一定的相似性[6], 本研究所测序列与锈红熊蜂的参考序列有3 bp(1个SNP和2个INDEL)差异, 然而与牧场熊蜂(No. DQ788077)、斯氏熊蜂(No. AF364828)、角熊蜂(No. DQ788121)以及黑足熊蜂(No. DQ787971)的参考序列之间却分别有27 bp(505/532)、14 bp(489/503)、57 bp(475/532)和54 bp(478/532)的差异。斯熊蜂尾部的浅橙色与部分密林熊蜂有一定的相似性[6], 本研究所测序列与斯熊蜂的参考序列有4 bp(全部为SNP)差异, 然而与密林熊蜂的参考序列(No. DQ788041)之间却有48 bp(488/536)的差异。可见, 这些形态上比较相似的物种, 在分子序列上出现混淆的可能性极低。因此, 基于形态特征比较, 同时参考分子序列信息, 可对这3个物种进行较为准确的鉴定。
从现有资料来看, 尽管对于青海省的熊蜂资源未有专门研究[19], 然而零星调查或馆藏记录的物种有38种之多[6, 7]。本研究显示, 青海东部地区有23种熊蜂, 占青海省现有物种记录的61%, 未发现有该省的新记录物种, 这可能与本研究区域较小有关。从亚属水平上看, 与其他地区相比, 青海东部地区的熊蜂(10个亚属)具有很高的多样性水平。例如, 中国北方地区8个省级行政单元(北京、天津、河北、山西、内蒙古、陕西、甘肃和宁夏)中, 只有甘肃省达到10个亚属[6]; 同时, 整个川渝地区也仅发现10个亚属[7]。在物种多样性方面, 目前只有甘肃[5]和河北[20]的研究提供了多样性数据。经过计算, 青海东部地区熊蜂的Shannon-Wiener多样性(2.67)介于甘肃(3.03)和河北(2.39)两省之间, 而其Pielou均匀度指数(0.85)则高于甘肃(0.76)和河北(0.71)两省。可见, 本研究区域的熊蜂物种多样性水平较高。
本研究发现, 不同物种之间的数量及其分布特征差异很大, 有3个物种(亚西伯熊蜂、红束熊蜂和克什米尔熊蜂)的数量都各自占总数的10%以上, 且在绝大多数采样点都有分布, 而有6个物种的数量占总体比例都低于1%, 且仅在一两个采样点出现(表2)。此外, 思壳拟熊蜂和牛拟熊蜂属于寄生型物种[7], 其数量非常有限。基于20个采样点(n=20)的Pearson相关性分析结果显示, 青海东部地区熊蜂物种多样性与环境因子之间有密切关联。物种丰富度随着经度的增加(越靠近东部)而增加, Shannon-Wiener多样性指数随年均温的升高而变大, Pielou均匀度指数则随着海拔的升高而降低。多元线性回归的结果与Pearson相关性分析结果基本一致。有趣的是, 西宁采样点的3个多样性指数都异常偏离线性回归曲线, 而去除这个采样点之后(n=19), 多样性指数与环境因子的关联度明显增强。去除西宁采样点后多元线性回归的结果显示, 3个多样性指数都与年均温关系最密切。此外, 无论是否去除西宁采样点的数据, 其他3个环境因子(纬度、年降水、年净初级生产力)都与物种多样性关系不大。综上, 与大多数动植物类群体现的规律一致[21], 气温是制约研究区域内熊蜂物种多样性的关键因子。另外, 有研究表明, 栖息地质量对熊蜂的生存至关重要, 人为活动可能严重威胁熊蜂物种多样性[22]。需要指出的是, 西宁采样点在西宁市南山, 处于城市边缘, 而其他采样点则全部位于偏僻的农区或牧区。本研究认为, 人为活动的干扰可能是导致西宁采样点3个多样性指标都偏低的主要原因。
自1985年德国科学家Roseler发现CO2麻醉可以打破熊蜂滞育以来[23], 熊蜂的周年繁育技术取得突破性进展。以荷兰科伯特公司(KOPPERT)为代表的熊蜂授粉服务公司对地熊蜂(B. terrestris)的授粉技术研发获得成功, 并在全球推广其商业品种。然而, 由于技术封锁和外来物种可能会导致生物入侵等问题[24, 25], 各国都在开发本土的熊蜂物种。目前, 美国、新西兰、日本等发达国家都分别成功开发出适合本国的优良熊蜂品种, 并大规模推广。中国的熊蜂研究起步较晚, 在吴杰[2]等科研人员的努力下取得了一系列成就, 对我国多种熊蜂成功进行周年繁育和小规模授粉试验。本研究采集到的物种中, 兰州熊蜂、密林熊蜂、明亮熊蜂、火红熊蜂已被证实具有市场开发潜力。其中, 明亮熊蜂群势较强, 授粉性能良好; 密林熊蜂群势较大(可达300~400头), 易于繁殖, 但容易受螟蛾类侵扰; 火红熊蜂具有采集植物种类多, 年活动时间长的优点[18]。最新的研究表明, 人工饲养条件下, 兰州熊蜂蜂王受精囊内精子数量、工蜂数量仅次于地熊蜂, 同时具有人工繁殖效率较高等优点[26]。笔者所在课题组的养殖试验显示, 黑尾熊蜂野生蜂王启动繁殖个体的比例与火红熊蜂相当。同时, 黑尾熊蜂还具有性格温顺、适应性强、生态幅宽的优点, 也具有一定的市场开发潜力。本研究将上述5种熊蜂合并, 作为可开发的经济物种, 并统计其在各采样点中的比例。结果显示, 总体上, 经济物种占总数的17.9%, 其中, 西宁、泽库、同德和乐都4个采样点的经济物种占比超过30%。另外, 将对这几个地区的熊蜂进行重点监测, 开展更为深入的生态学研究, 为青藏高原地区熊蜂资源的开发、利用和保护提供关键基础资料。
The authors have declared that no competing interests exist.
| [1] |
|
| [2] |
|
| [3] |
|
| [4] |
|
| [5] |
|
| [6] |
|
| [7] |
|
| [8] |
|
| [9] |
|
| [10] |
|
| [11] |
|
| [12] |
|
| [13] |
|
| [14] |
|
| [15] |
|
| [16] |
|
| [17] |
|
| [18] |
|
| [19] |
|
| [20] |
|
| [21] |
|
| [22] |
|
| [23] |
|
| [24] |
|
| [25] |
|
| [26] |
|