



{kind=link}
{kind=link}
{kind=link}
{kind=link}
高原鼠兔洞口密度对高山嵩草草甸土壤主要养分含量的影响
[张雯娜 , 金少红, 于成, 庞晓攀, 王娟, 郭正刚]
, 金少红, 于成, 庞晓攀, 王娟, 郭正刚]
, 金少红, 于成, 庞晓攀, 王娟, 郭正刚]
|
|


第一作者:张雯娜 (1995-),女,甘肃渭源人,在读硕士生,研究方向为草学。E-mail:[email protected]
高原鼠兔( Ochotona curzoniae)干扰会引起高山嵩草( Kobresia pygmaea)草甸土壤养分含量的变化。本研究分析了高原鼠兔干扰对高山嵩草草甸土壤主要养分的影响,并同步分析了高原鼠兔有效洞口密度和总洞口密度与土壤主要养分含量的关系。结果表明,高原鼠兔干扰显著增加了土壤0-20 cm土层有机碳、全氮、全磷、铵态氮、硝态氮含量( P<0.01),但显著降低了土壤速效磷含量;随着高原鼠兔有效洞口密度和总洞口密度增加,土壤0-20 cm土层全效养分(有机碳、全氮、全磷)含量先增加后降低,而速效养分(铵态氮、硝态氮、速效磷)含量先降低后增加,说明高原鼠兔总洞口密度和有效洞口密度与土壤养分含量的关系一致,因此当分析高原鼠兔干扰程度对土壤养分含量影响的趋势时,可采用总洞口密度代替有效洞口密度。
Disturbance in the population of the plateau pika, Ochotona curzoniae, often induces changes in the concentration of soil nutrients in alpine meadows. The effects of disturbance in the population of plateau pika on the concentration of the primary soil nutrients in a Kobresia pygmaea meadow was investigated to examine the relationships between the density of total burrow entrances and active burrow entrances of plateau pika and the concentration of the primary soil nutrients. Our results showed that the disturbance significantly increased the concentration of organic carbon (SOC), total nitrogen (TN), total phosphorus (TP), soil ammonium nitrogen (N$H_{4}^{+}$-N), and nitrate nitrogen (N$O_{3}^{+}$-N) in the soil, but there was a decrease in available phosphorus (AP) concentration in the 0-20 cm soil layer. As the densities of total burrow entrances and active burrow entrances increased, the total nutrient concentration in the 0-20 cm soil layer (SOC, TN, and TP) initially increased but then decreased. The nutrient concentration for N$H_{4}^{+}$-N, N$O_{3}^{+}$-N, and AP initially decreased then increased, indicating the relationship between the density of total burrow entrances and the concentration of soil nutrients are consistent with the density of active burrow entrances of plateau pika and the concentration of soil nutrients. Therefore, when determining the effects of plateau pika disturbances on the concentration of soil nutrients, the density of active burrow entrances can be replaced with the density of total burrow entrances.
土壤是高寒草甸植物繁殖和扩张的基床[1, 2], 而土壤养分是草原植物生长发育的基础, 更是维持植物群落生产力和稳定性的保障[2, 3, 4]。然而土壤养分含量往往易受生物干扰而改变, 甚至在短时间内发生重新配置[5], 其在生态环境脆弱、气候相对严寒的青藏高原地区尤为突出[6]。因此, 分析土壤养分含量对土壤扰动者的响应, 是维持青藏高原草地生态系统健康的重要内容。
高原鼠兔(Ochotona curzoniae)是青藏高原特有的营家族生活的小型啮齿类动物, 它们通过挖洞、覆土、采食、排泄和鼠尸等形式, 直接或间接地参与高寒草甸生态系统的能量流动和物质循环[7, 8], 因此被认为是高寒草甸生态系统中的一种主要的土壤生物干扰者[9]。目前关于高原鼠兔干扰对土壤的影响仍然存在争议。有研究表明, 高原鼠兔干扰对高寒草甸土壤会产生积极作用, 表现在增加土壤表层有机质含量、含水量和养分含量[7, 10]; 但另外一些研究却表明, 高原鼠兔干扰会对高寒草甸产生不利作用, 表现在降低土壤肥力, 增加CO2排放量[11], 降低优质牧草比例[12, 13], 加速高寒草甸退化[4, 14, 15]。造成这种差异的主要原因是有些研究考虑了高原鼠兔干扰强度[4, 15], 而有些研究则忽略了高原鼠兔的干扰强度[7, 10, 11, 12, 13, 14]。最新研究表明, 高原鼠兔干扰对高寒草甸土壤养分的影响不仅与干扰强度有关[4, 16], 还与草甸类型有关[17, 18]。
高原鼠兔具有移动特征, 且一年内能够生2~5胎[19], 因此利用高原鼠兔密度直接确定其干扰强度具有一定的难度。高原鼠兔对土壤的干扰, 是高原鼠兔多种行为的综合结果, 目前国内外均采用单位面积有效洞口密度划分高原鼠兔干扰强度[18, 20, 21]。然而高原鼠兔的废弃洞口, 依然能够通过影响土壤含水量和通气性而影响土壤性质, 表现在废弃鼠洞仍然能接纳天然降水入渗, 增加土壤含水量, 富集土壤有机质[7, 15]。陈剑等[22]认为高原鼠兔总洞口密度不仅能反映高原鼠兔干扰强度, 而且能反映高原鼠兔挖掘的活动量。事实上, 野外调查高原鼠兔洞口时, 测定总洞口密度要比测定有效洞口密度简单, 省时省力, 因此, 研究高原鼠兔干扰影响高寒草甸土壤性质时, 对究竟是总洞口密度还是有效洞口密度更具有指示性, 目前没有共识性结论。若有效洞口密度和总洞口密度两者之间可以相互替代, 那么用总洞口密度替代有效洞口密度, 将会简化调查工作的难度, 给野外调查工作带来极大便利, 节省人力和物力。然而高原鼠兔总洞口密度和有效洞口密度对高寒草甸土壤性质的影响是否趋同, 尚需科学研究提供证据。
青藏高原高寒草地中草甸类型较多[23], 其中高山嵩草(Kobresia pygmaea)草甸是高寒草地的主要类型之一, 也是高原鼠兔最适栖息的草甸类型之一[20]。本研究以高山嵩草草甸为对象, 通过分析高原鼠兔干扰对土壤主要养分的影响, 以及高原鼠兔总洞口密度和有效洞口密度与土壤养分含量的关系, 阐明能否用高原鼠兔总洞口密度替代有效洞口密度来研究高原鼠兔干扰对高寒草甸土壤性质的影响。
研究地区位于青藏高原东缘, 隶属甘肃省玛曲县境内(33° 06'-34° 30' N, 100° 40'-102° 29' E), 海拔3 300~4 806 m, 属典型高原大陆性气候, 多风、寒冷且潮湿, 年均温1.2 ℃, 月均温差异较大, 1月份的月均温低达-10 ℃, 7月份的月均温则高达11.7 ℃。年降水量约为564 mm, 主要集中在5-9月, 年蒸发量为1 000~1 500 mm。全年日照时数约为2 613.9 h, 年内相对无霜期仅19 d, 没有绝对无霜期。
土壤类型为亚高山草甸土, 其中营养物质多以有机态形式存在, 表层草皮层约有7 cm厚, 有机质含量可达到100~150 g· kg-1, 土壤潜在肥力较高[24]。草地类型为高寒草甸, 以莎草科嵩草属的高山嵩草为主要优势种, 常见伴生种有垂穗披碱草(Elymus nutans)、小花草玉梅(Anemone rivularis var. flore-minore)、钝裂银莲花(Anemone obtusiloba)、莓叶委陵菜(Potentilla fragarioides)等[25, 26, 27]。
2016年7月在玛曲县完成野外样地调查和取样。调查区为围栏的冷季草地, 即夏季不放牧, 围栏主要是避免夏季其他草食动物的干扰, 且便于冷季家畜放牧。首先选择高原鼠兔干扰样地, 以高原鼠兔洞口和高原鼠兔出没为基准。然后根据草甸类型和地貌特征, 在干扰样地附近选择等同面积的未干扰样地, 以没有发现高原鼠兔和高原鼠兔洞口为基准[28]。驱车沿公路前行下一个地点, 选择另外一个干扰样地和未干扰样地, 共计选择10个干扰样地和10个未干扰样地。10个干扰样地和10个未干扰样地地貌基本一致, 草甸类型均为高山嵩草草甸, 植物群落组分基本相似, 优势种为高山嵩草, 主要伴生种有达乌里秦艽(Gentiana dahurica)、小花草玉梅、莓叶委陵菜等[25, 26, 27]。各研究地点之间的距离为1~5 km, 每个地点干扰样地和未干扰样地之间距离为500~1 000 m, 形成一个缓冲区域, 干扰样地和未干扰样地整体上呈现为交错分布状态[17, 29]。样地面积为25 m× 25 m, 每个样地的四角采用木桩标记, 四周采用塑料绳标记。干扰样地内, 观测高原鼠兔的总洞口数(包括新鲜、废弃及枯竭的洞穴)并记录[29], 然后采用3 d堵洞法测定各样地的有效洞口数并取其平均值[20, 30, 31], 10个干扰样地内有效洞口密度和总洞口密度变化较大(表1)。在各干扰样地和未干扰样地内采用“ W” 型布置5个1 m× 1 m的样方, 样方间隔约8 m, 在每个样方内用直径为10 cm的土钻随机收集0-20 cm的土样3次, 在干扰样地内避免取样点位于洞口和裸斑处, 将3次取得的土样混合均匀成一个土样, 总共收集到100个土样, 将所有土样均装入密封袋带回实验室分析。
| 表1 样地内高原鼠兔洞口密度 Table 1 The densities of burrow entrances for plateau pika in study areas entrance· hm-2 |
实验室内首先去除土壤样品中的植物调落物, 根系及石头等杂质, 然后在室温条件下自然风干, 再经充分研磨后过0.25 mm的尼龙筛, 以分析土壤N、P、K和C含量。土壤全氮(total nitroges, TN)采用凯氏定氮法测定, 土壤有机碳(soil organic carbon, SOC)采用重铬酸钾容量法-外加热法测定, 土壤铵态氮(N
首先采用独立样本T检验分析高原鼠兔干扰对土壤氮、磷、钾和碳含量的影响是否显著, 若差异不显著, 则不做进一步分析; 若差异显著, 再用R-3.3.2-windows软件的ggplot2程序包拟合干扰梯度和土壤养分含量的关系。
高原鼠兔干扰显著影响了土壤有机碳(F=1.034, df=18, sig(双侧)=0.032)、全氮(F=0.043, df=18, sig(双侧)=0.003)、全磷(F=1.744, df=18, sig(双侧)=0.038)、铵态氮(F=0.012, df=18, sig(双侧)=0.027)、硝态氮(F=0.814, df=18, sig(双侧)=0.030)和速效磷(F=0.214, df=18, sig(双侧)=0.047)含量, 但土壤全钾和速效钾含量的影响不显著(P> 0.05), 表现为干扰样地土壤有机碳、全氮、全磷、铵态氮和硝态氮含量均显著高于未干扰样地的土壤有机碳、全氮、全磷、铵态氮和硝态氮含量(P< 0.05), 而干扰样地土壤速效磷含量却显著低于未干扰样地土壤速效磷含量(图1)。
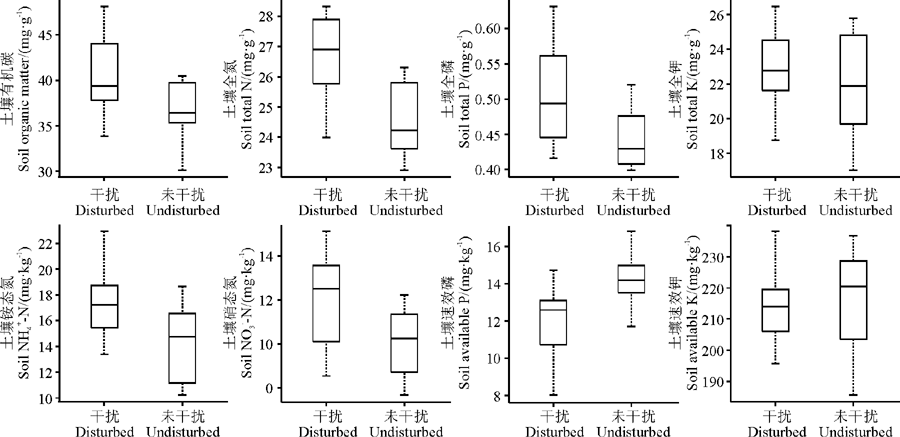 | 图1 高原鼠兔干扰对土壤养分含量的影响Fig. 1 Effects of plateau pika disturbance on the concentration of soil nutrients |
高原鼠兔有效洞口密度和总洞口密度与土壤有机碳含量的关系基本趋同, 表现为随高原鼠兔有效洞口密度和总洞口密度增加, 土壤有机碳含量先增加后降低, 分别在670和1 271个· hm-2时最高(图2)。
 | 图2 高原鼠兔洞口密度对土壤有机碳含量的影响图中灰色阴影部分表示95%的置信区间, 垂直线表示拐点位置, ABED表示有效洞口密度, TBED表示总洞口密度, 下同。Fig. 2 Effects of the density of burrow entrances of plateau pika on the concentration of organic carbon in the soilThe shaded part of the figure indicates the 95% confidence interval, the vertical line indicates the position of the inflection point, ABED indicates the density of active burrow entrances, TBED indicates the density of total burrow entrances; similarly for the following figures. |
随高原鼠兔有效洞口密度和总洞口密度增加, 土壤全氮和速效氮含量变化趋势不一致, 其中土壤全氮含量先增加后降低, 在754和1 381个· hm-2时达到最高, 然而土壤铵态氮和硝态氮含量却先降低后增加, 其中铵态氮含量在有效洞口密度为717 个· hm-2, 总洞口密度为1 345个· hm-2时最低, 硝态氮含量在有效洞口密度为577个· hm-2, 总洞口密度为1 156个· hm-2时最低(图3)。
 | 图3 高原鼠兔洞口密度对土壤氮含量的影响Fig. 3 Effects of the density of burrow entrances of plateau pika on the concentration of nitrogen in the soil |
高原鼠兔有效洞口密度和总洞口密度对土壤全磷含量和速效磷含量的影响不同。随高原鼠兔有效洞口密度和总洞口密度增加, 土壤全磷含量表现为先增加后降低, 且在有效洞口密度为666个· hm-2, 总洞口密度为1 264个· hm-2时达到最高; 而土壤速效磷含量却表现为先降低后增加, 在有效洞口密度为697个· hm-2, 总洞口密度为1 306个· hm-2时最低(图4)。
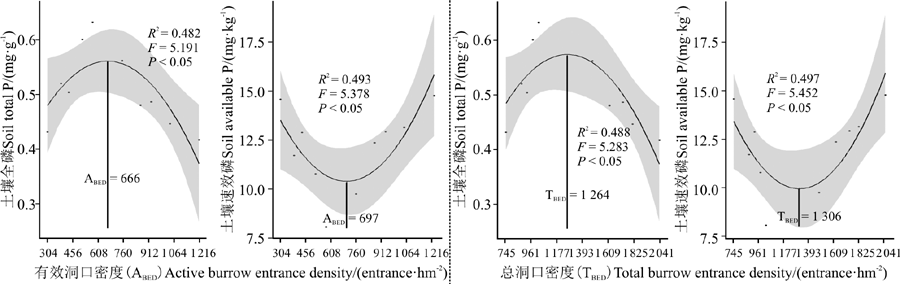 | 图4 高原鼠兔洞口密度对土壤磷含量的影响Fig. 4 Effects of the density of burrow entrances of plateau pika on the concentration of phosphorus in the soil |
作为土壤扰动者, 高原鼠兔是高寒草甸土壤养分含量变化的主要影响者之一[33, 34]。本研究表明, 高原鼠兔干扰虽然没有显著影响土壤全钾和速效钾含量, 但显著提高了土壤有机碳、全氮、全磷、铵态氮和硝态氮含量, 降低了土壤速效磷含量, 这与囊鼠干扰增加北美草原土壤全氮含量[35], 土拨鼠干扰增加奇瓦瓦州草原土壤碳储量的研究结果一致[36]。高原鼠兔干扰增加有机碳和全氮含量的主要途径可能包括下列几个方面:首先, 高原鼠兔覆土作用掩埋植被[14, 37], 增加了土壤有机碳的输入量; 其次, 高原鼠兔排泄物会形成肥岛[34, 38], 增加土壤有机质输入量, 因此, 高原鼠兔干扰样地内土壤有机碳和全氮含量增加。土壤铵态氮和硝态氮含量取决于土壤有机质矿化速率和参与分解的微生物的活性[7, 39], 高原鼠兔干扰后土壤有机碳和全氮含量的增加会提高土壤微生物获得自身氮素的能力, 从而间接提高了有机质的分解速率[10], 加快土壤矿化作用[33]; 然而高原鼠兔干扰加速了土壤水分从表层向深层的渗透[17], 其淋溶作用使土壤铵态氮和硝态氮向深层土壤转移, 另外, 植物吸收一定量的铵态氮和硝态氮, 但整体上铵态氮和硝态氮减少量可能小于矿化作用增加的量, 因此高原鼠兔干扰最终会增加土壤浅层的铵态氮和硝态氮含量。土壤全磷和全钾含量主要取决于土壤母质, 而土壤全磷包含有机磷和无机磷, 有机磷含量与土壤有机质含量正相关, 而土壤有机质中含钾量较少[40], 因此, 高原鼠兔干扰对土壤钾含量的变化影响甚微, 出现母质相同时, 高原鼠兔干扰会显著增加土壤全磷含量, 而对土壤全钾含量没有影响的现象。高原鼠兔干扰降低了土壤速效磷含量, 主要是由于高原鼠兔干扰后土壤水分下渗能力增加, 淋溶作用将速效磷带到深层土壤, 同时植物对养分的吸收作用增强[17], 从而导致浅层土壤速效磷降低。土壤速效钾含量主要取决于土壤阳离子交换量(CEC)[40], 这主要取决于土壤类型, 由于干扰样地和未干扰样地土壤类型一致, 故高原鼠兔干扰没有明显影响土壤速效钾含量。
高原鼠兔有效洞口密度和总洞口密度对土壤主要养分含量的影响趋势一致, 其中全效养分含量(土壤有机碳、全氮和全磷)随高原鼠兔有效洞口密度和总洞口密度增加呈现先增加后降低的变化过程, 而速效养分含量(铵态氮, 硝态氮, 速效磷)呈现为先降低后增加的变化趋势, 这与Pang和Guo[18]以及Yu等[17]对高原鼠兔有效洞口密度与土壤主要养分含量关系的研究结果类似, 说明高原鼠兔干扰对土壤养分的影响存在理论阈值, 且高原鼠兔总洞口密度的阈值大于有效洞口密度, 干扰程度在有效洞口密度与总洞口密度各自的阈值范围内, 土壤全效养分含量均会显著增加, 而干扰强度低于或高于该阈值时, 全效养分含量会降低[17, 18, 41]。在低干扰强度时, 草地植被盖度和生物量相对较高, 植被缓冲作用使土壤温度较低[16], 且致密草毡层使土壤水分下渗困难, 以地表径流形式流失[15], 土壤含水量较低, 因此微生物活性变化不大, 有机质分解作用和土壤矿化作用变化不大。与低干扰强度相比, 中等干扰强度时高原鼠兔能够掩埋更多的植被, 尿液和粪便等排泄物也会增多, 土壤有机质和全氮输入增加, 适宜高原鼠兔干扰增加了土壤通透性和持水能力[16, 24], 土壤温度相对升高, 矿化作用和分解作用相对增强, 但整体上表现为输入大于输出, 因此中干扰强度时土壤有机质和全氮含量达到最大; 当高原鼠兔干扰强度超过阈值时, 植被盖度和生物量更低[42, 43], 土壤有机质的来源大幅度减少, 高原鼠兔挖掘活动产生的裸斑面积增大, 土壤温度和通气性增强, 蒸发量增大[18, 21], 从而刺激微生物活性增强, 促进了有机质的分解速率, 土壤矿化作用增强, 使土壤有机质和全氮输出增加, 因此土壤有机碳和全氮含量降低。土壤铵态氮、硝态氮和速效磷含量在高原鼠兔有效洞口密度与总洞口密度阈值范围内显著降低, 主要原因可能是中等干扰程度下, 植被盖度和生物量虽然有一定程度的降低[25], 但植物群落组分内禾本科植物比例或类禾草植物比例显著增加, 而禾本科植物喜食氮素[18], 从而使铵态氮和硝态氮含量降低, 随干扰强度增加, 植物组分杂草比例增加[28], 加之植被盖度和生物量大幅度降低[42, 43], 其吸收和利用铵态氮和硝态氮的能力减弱, 使铵态氮和硝态氮含量逐渐增加。土壤中速效磷主要来源于土壤母质, 高原鼠兔干扰后在淋溶作用的影响下, 速效磷含量逐渐降低, 同时在高原鼠兔中度干扰时, 其洞口附近的植物生物量有所改善[14], 植物对养分的吸收作用增强[17], 因此在中度干扰时土壤速效磷含量最低。随高原鼠兔干扰的增加植被盖度和生物量显著降低[42, 43], 植物吸收和利用速效磷的能力减弱, 淋溶作用降低的量可能小于植被吸收利用减少而增加的量, 因此速效磷含量逐渐升高。
综上, 高原鼠兔干扰对土壤全效养分和速效养分含量的影响不一致, 高原鼠兔有效洞口密度和总洞口密度与土壤养分含量的关系一致, 但采用总洞口密度测定时土壤养分含量变化的拐点大于有效洞口密度, 二者变化过程趋同。因此, 在评估高原鼠兔干扰程度对高寒草甸土壤养分含量影响的趋势时, 可考虑用研究区域内高原鼠兔总洞口密度代替有效洞口密度, 可以改进有效洞口密度测定时费工费时费力的不足之处。
(责任编辑 武艳培)
The authors have declared that no competing interests exist.
| [1] |
|
| [2] |
|
| [3] |
|
| [4] |
|
| [5] |
|
| [6] |
|
| [7] |
|
| [8] |
|
| [9] |
|
| [10] |
|
| [11] |
|
| [12] |
|
| [13] |
|
| [14] |
|
| [15] |
|
| [16] |
|
| [17] |
|
| [18] |
|
| [19] |
|
| [20] |
|
| [21] |
|
| [22] |
|
| [23] |
|
| [24] |
|
| [25] |
|
| [26] |
|
| [27] |
|
| [28] |
|
| [29] |
|
| [30] |
|
| [31] |
|
| [32] |
|
| [33] |
|
| [34] |
|
| [35] |
|
| [36] |
|
| [37] |
|
| [38] |
|
| [39] |
|
| [40] |
|
| [41] |
|
| [42] |
|
| [43] |
|