



{kind=link}
{kind=link}
{kind=link}
{kind=link}
几种环境因子对黄花矶松种子萌发的影响
[王靖靖1  , 余玲
, 余玲1 , 朱恭2 , 陈香来1 , 胡小文1 ]
, 余玲]
|
|


第一作者:王靖靖(1994-),女,河南灵宝人,在读硕士生,研究方向为种子生态学。E-mail:wangjj2017@lzu.edu.cn
以黄花矶松( Limonium aureum)种子为材料,通过培养皿纸上发芽法和盆钵法,研究了温度、光照、水分、盐分和埋藏深度几种环境因子对黄花矶松种子萌发的影响。结果表明,1)温度、光照及其互作显著影响种子萌发( P<0.001),随温度升高,种子萌发率与萌发速率先升高后降低,在温度为25、30和20/30 ℃时最高;在35和40 ℃时,光照显著促进种子萌发( P<0.05),其他温度条件下无显著效应( P>0.05);2)种子萌发率与萌发速率随干旱胁迫和盐胁迫程度的增加呈下降趋势,对水分与盐分胁迫耐受的半致死浓度分别为-0.64和-0.84 MPa(0.201 mol·L-1),在低渗透势下,种子在盐分胁迫下比水分胁迫下萌发更好;3)随埋藏深度增加,黄花矶松种子萌发率呈现先增加后减少的趋势,其适宜播种深度为0.5 cm。
The effect of temperature, light, water, salt, and burial depth on seed germination of Limonium aureum was analyzed. The results indicated that: 1) Temperature, light and their interaction significantly affected L. aureum seed germination ( P<0.001). Germination percentage and germination rate increased initially and then decreased with increasing temperature, and reached the highest when the temperature was 25, 30, and 20/30 ℃. When the temperature was 35 and 40 ℃, light promoted seed germination of L. aureum significantly ( P<0.05), whereas light showed no effect at other temperatures ( P>0.05); 2) With increasing drought and salt stress, germination percentage and germination rate of L. aureum decreased gradually. The median lethal concentrations of drought resistance and salt tolerance for L. aureum were -0.64 and -0.84 MPa (0.201 mol·L-1), respectively. When osmotic potential was low, seeds under salt stress germinated better than those under drought stress; 3) The optimal sowing depth of L. aureum seeds was 0.5 cm.; therefore, these seeds are possibly suitable for shallow seedling.
黄花矶松(Limonium aureum)又名黄花补血草、金色补血草、金匙草等, 白花丹科(Plumbaginaceae)补血草属(Limonium), 多年生旱生泌盐草本植物[1]。其具有抗盐碱、抗干旱等特性, 对恶劣环境的适应性极强, 常见于滩地、流动沙丘、戈壁、荒漠等干旱半干旱荒漠地区。在我国呼伦贝尔沙地、河西走廊沙地以及华北北部等地均有分布[2, 3]。作为干旱荒漠地区为数不多的野生花卉之一, 黄花矶松有着十分重要的观赏、生态和药用等价值。
黄花矶松在恶劣环境下能够生长发育的前提是种子能够萌发。然而, 种子萌发通常是植物生活史最为脆弱的阶段, 容易受外界环境因子如温度、水分、光照等影响[4, 5]。温度是影响植物时空分布的控制因子, 对种子萌发以及出苗起到关键性作用, 能够促进或抑制种子吸水, 影响酶促过程[6, 7]; 光照作为一种信号分子影响种子萌发, 促进或抑制种子休眠, 进而调控种子在适宜条件下的萌发[8, 9]; 水分是干旱半干旱地区种子萌发的决定因素, 尤其是荒漠盐渍化地区, 主要通过渗透胁迫影响种子的萌发[10]; 盐分是盐渍化地区种子萌发的限制因子, 严重影响该地区种子的萌发以及出苗生长; 埋藏深度直接影响植物的出苗率以及出苗速率[11]。因此, 研究多环境因子对种子萌发的影响对于明确黄花矶松在自然条件下的萌发机制、栽培利用等都具有重要意义。
目前, 围绕黄花矶松引种栽培、组织培养、育苗技术、化学成分、超干保存、结构解剖学以及盐胁迫机制等方面的研究工作报道较多[12, 13], 但有关其种子萌发特性的研究比较零散, 如李阳春等[14]和王俊年等[15]研究了温度对其种子萌发的影响; 高天鹏等[16]和马全林等[17]探究了埋深对其种子萌发的作用; 尤佳等[18, 19]探讨了盐胁迫对其种子萌发的影响机理。这些研究都从单一方面为探究黄花矶松种子的萌发特性提供了参考。但种子萌发通常会受到多种环境因子的影响, 因此系统研究多种环境因子对黄花矶松种子萌发、出苗以及生理生态学特性具重要意义。此外, 黄花矶松作为一种旱生泌盐植物, 目前尚不清晰其对干旱和盐渍化的耐受程度, 所以开展黄花矶松种子萌发对水分和盐分胁迫的耐受阈值分析研究, 可为明确黄花矶松种子抗旱耐盐性能, 发掘其在荒漠盐渍化地区土壤改良方面的潜能奠定基础。
本研究于2017年3月至2017年6月在兰州大学草地农业科技学院榆中校区实验站完成。萌发试验于光照培养箱中进行, 埋深试验于智能温室中进行。
供试种为野生种, 于2016年采集于甘肃省民勤县沙生植物园, 种子清选后贮藏于室温条件下备用。千粒重为(0.73± 0.05) g。
1.3.1 温度和光照 采用培养皿纸上发芽法, 设置5、10、15、20、25、30、35、40、45 ℃共9个恒温和15/25、20/30 ℃两个变温条件以及12 h光照/12 h黑暗(12 h L/12 h D)、24 h黑暗(24 h D)两个光照条件。试验中使用双层锡箔纸将培养皿包裹严实来模拟全黑暗条件。每个处理设置4个重复, 每个重复50粒种子。以胚根突破种皮为标准来判断种子是否萌发, 萌发高峰期增加统计次数, 其余时间每24 h统计一次, 种子萌发高峰期过后连续3 d没有种子萌发即视为试验结束, 试验期共21 d。全黑暗条件下的样品仅于试验结束后统计一次, 以防试验期内统计时遇光影响结果的可靠性, 与光照条件下的样品同时结束, 以确保可比较性。
1.3.2 水分胁迫和NaCl胁迫 同上使用培养皿纸上发芽法, 在25 ℃、12 h L/12 h D条件下, 根据Michel和Kaufmann[20]的方法按照公式计算设置水势为0(CK)、-0.3、-0.6、-0.9、-1.2和-1.5 MPa的6个PEG-6000溶液以及渗透势为0(CK)、-0.3、-0.6、-0.9、-1.2、-1.5、-1.8、-2.1和-2.4 MPa的9个NaCl溶液, NaCl溶液对应的摩尔浓度依次为0、0.090、0.151、0.213、0.274、0.336、0.398、0.459、0.521 mol· L-1。每个处理设置4个重复, 每个重复50粒种子, 萌发标准同上。每24 h统计一次, 种子萌发高峰期过后连续3 d没有种子萌发即视为试验结束, 胁迫期共14 d。在水分胁迫和盐胁迫试验结束后立即进行恢复试验, 即将胁迫处理条件下未萌发的种子转移到加有蒸馏水的培养皿中, 定期统计萌发个数直至试验结束并且计算恢复萌发率, 恢复试验结束标准同上, 恢复期为11 d。
1.3.3 埋深 埋深试验采用盆钵法, 设置8个埋深梯度:0(CK)、0.25、0.5、1.0、1.5、2.0、3.0和4.0 cm, 每个处理设3个重复, 每个重复100粒黄花矶松种子。基质沙层和覆盖沙层购买于榆中县夏官营镇沙场, 均需过筛(粒径2 mm)且130 ℃高温消毒2 h。黄花矶松子叶突出表层即视为出苗, 试验期间每24 h观察一次出苗情况, 种子出苗高峰期过后连续3 d没有种子出苗即视为试验结束, 试验周期21 d。
共测定萌发率、出苗率、萌发指数、出苗指数、恢复萌发率(recovering germination percentage, RGP)[21]、总萌发率(total germination percentage, TGP)[21]、耐旱(盐)适宜浓度、耐旱(盐)半致死浓度、耐旱(盐)完全致死浓度。
萌发(出苗)率=(n/N)× 100%;
式中:n为供试种子中萌发(出苗)的种子数; N为供试种子数。
萌发(出苗)指数=∑ (Gt/Dt);
式中:Gt为供试种子中与Dt相对应每天萌发(出苗)的种子数; Dt为萌发天数。
RGP=[m/(N-n)]× 100%;
式中:m为恢复试验中萌发的种子数; n为胁迫试验中萌发的种子数; N为进行胁迫试验的总种子数。
TGP=[(m+n)/N]× 100%;
式中:m为恢复试验中萌发的种子数; n为胁迫试验中萌发的种子数; N为进行胁迫试验的总种子数。
耐旱(盐)适宜浓度:种子萌发率为对照的75%时所对应的渗透势。
耐旱(盐)半致死浓度:种子萌发率为对照的50%时所对应的渗透势。
耐旱(盐)完全致死浓度:种子萌发率为0时所对应的渗透势。
数据采用Excel 2013录入并作图, SPSS 21.0进行统计分析, 用Duncan法进行One-way(P< 0.05)、Two-way ANOVA分析(P< 0.001)和线性回归分析(P< 0.001), 用LSD多重比较法检验其差异显著性。
随温度的升高黄花矶松种子萌发率以及萌发指数均呈现先升高后降低的趋势(图1)。萌发率在大于30 ℃的高温以及低于15 ℃时显著下降(P< 0.05)。萌发指数在25、30和20/30 ℃时较高, 相比于20~30 ℃, 5~15和35~45 ℃温度条件下萌发指数均显著下降(P< 0.05)(图1)。
 | 图1 温度和光照对黄花矶松种子萌发率和萌发指数的影响 同一因素不同处理间不同小写字母表示差异显著(P< 0.05), 下同。Fig. 1 Effect of temperature and light on the germination percentage and index of Limonium aureum Different lowercase letters represent significant differences aming the different treatments under the same factor at the 0.05 level; similarly for the following figures. |
35和40 ℃时, 光照条件下黄花矶松种子萌发率显著高于黑暗条件下(P< 0.05)(图1), 其余温度条件下有无光照不影响黄花矶松种子的萌发率(P> 0.05)。
双因素方差分析的结果表明, 温度、光照以及二者互作均极显著影响黄花矶松种子的萌发率(P< 0.001)(表1)。
| 表1 温度、光照及其互作对黄花矶松种子萌发率的影响的双因素方差分析 Table 1 Two-way ANOVA of the effect of temperature, light and their interaction on the germination percentage of Limonium aureum |
随着水分胁迫和盐胁迫的加重, 黄花矶松种子的萌发率和萌发指数逐渐降低(图2)。当PEG溶液渗透势为-1.5 MPa时无种子萌发。然而, 当NaCl溶液渗透势为-2.4 MPa时仍有8%的种子能够萌发。
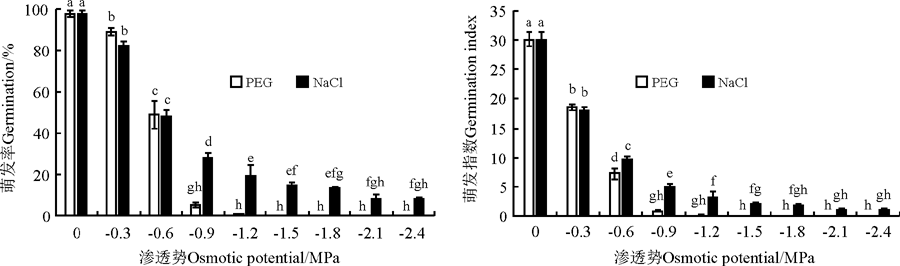 | 图2 水分胁迫和NaCl胁迫对黄花矶松种子萌发率和萌发指数的影响Fig. 2 Effect of water and NaCl stress on germination percentage and index of Limonium aureum |
在等渗的PEG和NaCl条件下, 黄花矶松种子除了萌发率在0、-0.3和-0.6 MPa以及萌发指数在0和-0.3 MPa无显著差异(P> 0.05)(图2)外, 在其他渗透势下, 盐胁迫条件下的萌发率和萌发指数均显著高于水分胁迫条件下(P< 0.05)。
回归分析结果表明, 无论是干旱胁迫还是盐胁迫, 黄花矶松种子萌发率和渗透势均呈极显著正相关关系(P< 0.001)(表2), 胁迫对种子萌发率的影响程度为PEG> NaCl, 说明PEG胁迫对黄花矶松种子萌发的抑制作用更强。在这两种等渗溶液中, 种子萌发的抗旱和耐盐半致死浓度分别为-0.64和-0.84 MPa。黄花矶松种子萌发对两种胁迫的耐受程度为适宜浓度是PEG> NaCl; 而半致死浓度值和和完全致死浓度是NaCl> PEG。
| 表2 黄花矶松种子萌发率与渗透势的关系 Table 2 The relationship between the germination percentage of Limonium aureum and osmotic potential |
随PEG溶液和NaCl溶液渗透势的下降, 黄花矶松种子的恢复萌发率逐渐升高, 而总萌发率呈现先降低后升高的趋势, 在-0.6 MPa时最小(图3)。在等渗的PEG和NaCl胁迫条件下, 黄花矶松种子恢复萌发率只有在-0.3 MPa以及总萌发率在-0.6 MPa时, NaCl显著高于等渗的PEG(P< 0.05), 在其余条件下均无显著差异(P> 0.05)。
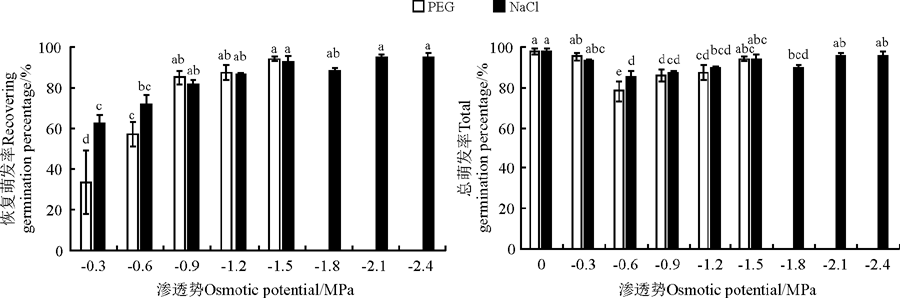 | 图3 水分胁迫和NaCl胁迫对黄花矶松种子恢复萌发率和总萌发率的影响Fig. 3 Effect of water and NaCl stress on recovering germination percentage and total germination percentage of Limonium aureum |
随埋深增加黄花矶松出苗率和出苗指数均呈现先升高后下降的趋势(图4)。埋深0.5 cm 时, 出苗率与出苗速率达到最大, 而后随埋深增加, 出苗率显著下降(P< 0.05), 埋深3 cm及以上时, 出苗率为0。
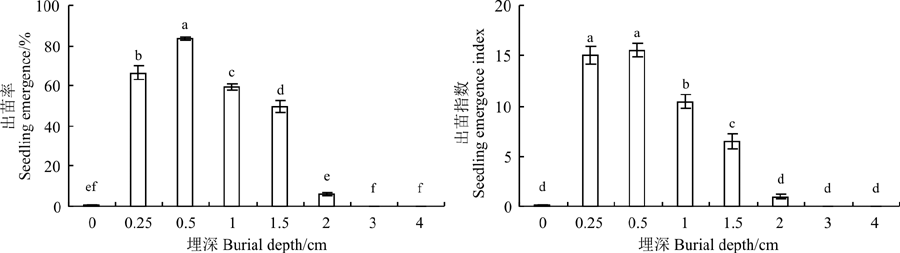 | 图4 埋藏深度对黄花矶松种子出苗率和出苗指数的影响Fig. 4 Effect of buried depth on seeding emergence percentage and seeding emergence index of Limonium aureum |
温度是植物生长发育过程中一个至关重要的环境因子[22]。温度过低或过高都不利于种子的萌发和生长, 如温度太低, 酶活性过低不足以催化各种代谢反应进行, 而温度太高导致酶变性失活[23, 24]。本研究表明, 黄花矶松种子在5~45 ℃均能萌发, 表明其具有较宽的萌发温幅。这与李阳春和刘存琦[14]的研究结果一致, 其研究发现黄花矶松种子在5~40 ℃均可萌发。此外, 黄花矶松在10~40 ℃条件下萌发率均超过50%, 说明其适宜在较大温度范围内萌发建植。随温度升高萌发率和萌发指数均表现为先升高后降低的趋势, 其最适温度为恒温25和30 ℃以及变温20/30 ℃, 这与王俊年和李得禄[15]的研究结果(15~20 ℃)有异议, 两个研究的种子均是采自甘肃省民勤县沙生植物园, 生境相同, 但是王俊年和李得禄[15]的试验材料是当年秋季采集, 于10月进行萌发试验, 为新鲜种子; 而本研究所用种子经过半年时间贮藏。可能随着贮藏时间的增加, 种子会缓慢后熟, 休眠释放, 进而改变种子的萌发行为[25, 26]。
光照和温度一样, 亦是影响种子萌发和幼苗建成的重要环境因子。植物种子萌发究竟是否需要光照的存在, 这不仅由植物自身遗传物质决定, 还受到外部环境因子的影响[27]。本研究表明, 光照对种子萌发的影响因温度而异, 如黄花矶松种子在温度为35和40 ℃的高温时, 光照显著促进了其种子的萌发率, 而在其余温度条件下有无光照并不影响黄花矶松种子的萌发率。说明35和40 ℃的高温会诱导黄花矶松种子形成次生休眠[4, 7, 8], 而光照可以作为一种信号分子抑制休眠[25, 28], 因此, 高温条件下光照的存在会促进种子萌发。
植物种子对水分胁迫的响应一定程度反映着其抗旱性强弱, 而抗旱性强弱代表了植物在干旱环境中的生存能力[29]。随着溶液渗透势逐渐降低, 黄花矶松种子的萌发率和萌发指数也逐渐降低, 这与刘萍等[30]对补血草(Limonium)种子的研究结果一致; 黄花矶松种子在渗透势为-1.5 MPa时种子萌发完全受到抑制, 而耳叶补血草(L. otolepis)和簇枝补血草(L. chrysocomum)种子在渗透势为-0.9 MPa时萌发完全受到抑制[30, 31], 表明黄花矶松种子的抗旱性强于耳叶补血草和簇枝补血草。黄花矶松种子的抗旱半致死浓度和完全致死浓度分别为-0.64和-1.28 MPa, 大于盐节木(Halocnermum strobilaceum)种子的抗旱半致死浓度(-0.75 MPa)和完全致死浓度(-1.46 MPa)[32], 说明盐节木更抗旱, 这可能与盐节木是矮小半灌木这一特性有关。
盐分对植物种子萌发的影响程度决定着该植物对盐渍化土壤的适应度, 盐溶液通过渗透胁迫阻碍种子吸水和(或)盐离子效应造成离子毒害进而抑制种子萌发[18, 33]。黄花矶松种子萌发率和萌发指数均随着NaCl胁迫的加重而逐渐下降, 这和尤佳等[18, 19]研究结果一致。当NaCl浓度为0.521 mol· L-1(-2.4 MPa)时, 黄花矶松萌发率为8%。而耳叶补血草和簇枝补血草种子在NaCl浓度为0.4 mol· L-1时萌发已完全受到抑制, 说明黄花矶松的耐盐性要强于另外两种补血草[30, 31]。此外, 黄花矶松种子萌发期的耐盐半致死浓度和完全致死浓度分别为0.201 mol· L-1(-0.84 MPa)和0.471 mol· L-1(-2.16 MPa), 小于盐节木种子的耐盐半致死浓度(0.47 mol· L-1)和完全致死浓度(0.83 mol· L-1)[32], 这表明盐节木种子相比于黄花矶松种子而言更具耐盐能力。但与同为补血草属植物的二色补血草(L. bicolor)以及甜土植物水稻(Oryza sativa)、大豆(Glycine max)相比, 黄花矶松种子的耐盐半致死浓度大于二色补血草、水稻和大豆的耐盐半致死浓度[34, 35]。
有研究表明, 同一植物对等渗的水分胁迫和盐胁迫的敏感程度不同[32]。本研究中, 当渗透势低于-0.6 MPa时黄花矶松种子在盐胁迫条件下的萌发率和萌发指数基本都要高于等渗的水分胁迫条件, 并且当NaCl渗透势为-2.4 MPa时仍然有8%的萌发率。说明盐胁迫不利于黄花矶松种子萌发主要是由渗透胁迫造成的, 而离子毒害作用较小甚至没有。究其原因, 可能是因为黄花矶松为盐生植物, 少量Na+低浓度盐可以促进渗透调节、降低种子渗透势、加速种子吸水和刺激呼吸酶产生作用, 而PEG为大分子物质, 不易进入种子内部[18, 36, 37]。
随水分胁迫和盐胁迫的加重, 黄花矶松种子的恢复萌发率逐渐升高, 高胁迫条件下的总萌发率与对照无显著差异。这表明高强度的干旱胁迫和盐胁迫并没有对黄花矶松种子造成不可逆的损害, 没有使种子永久性的丧失萌发能力, 仅是暂时地抑制了种子吸水[29, 38], 种子仍保持较高的萌发潜力。一旦胁迫解除进行复水萌发, 黄花矶松种子会恢复其萌发能力, 达到较高的萌发率。这可能是黄花矶松种子能够在盐碱地以及干旱地区萌发的关键原因。此外, 黄花矶松的总萌发率随着两种胁迫程度的加深呈先降低后升高的趋势, 在中等渗透势条件下最低。可能是由于在中等渗透势条件下诱导了黄花矶松种子形成生理休眠, 萌发潜力降低, 使得恢复萌发率较低, 进而总萌发率最低[39]。有研究表明, 盐生植物主要有耐盐型和避盐型两种生存策略。耐盐型植物在高盐条件下主要表现为高萌发率和(或)高萌发速率, 而避盐型植物表现为高恢复萌发率[21]。黄花矶松在高盐条件下有高恢复萌发率, 表明黄花矶松为避盐型植物。恢复研究表明, 黄花矶松种子总萌发率都有所增加, 这不仅体现了黄花矶松耐旱耐盐这一大特性, 也反映了黄花矶松长期生存于荒漠干旱环境中, 种子萌发持续而分散、萌发速率低, 是其在与干旱环境千百万年的博弈中而演化出的萌发适应对策[40, 41]。这种策略提高了黄花矶松种子的利用率, 同时由于其萌发时间的不一致性, 降低了因外界环境巨变造成物种灭绝的风险。
研究结果显示, 随着埋藏深度的增加黄花矶松出苗率和出苗速率先升高后下降。黄花矶松种子最适埋藏深度为0.5 cm, 这与高天鹏等[16]和马全林等[17]研究结果一致。当种子置于基质表层时, 出苗率比较低可能是由于表层水分蒸发过快, 使得水分不能满足种子出苗所需。当埋藏深度达到3和4 cm时没有种子出苗, 可能是由于以下原因造成:1)随着埋藏深度增加, 顶土压力增大, 而黄花矶松种子较小, 种子自身能量不足以让其突破阻碍到达表面[11, 42]; 2)当埋深到达一定程度时, 土壤中含氧量满足不了种子萌发的氧分压, 种子进行无氧呼吸, 呼吸产物对种子产生毒害作用, 代谢气体与周围气体环境之间的相互作用导致了种子休眠或死亡[42], 故黄花矶松种子适合浅播。
综上所述:1)黄花矶松种子的最适萌发范围为25、30和20/30 ℃; 2)当温度为35和40 ℃时光照的存在可以更好地促进黄花矶松种子萌发, 光照对黄花矶松种子萌发的影响依赖于温度的变化; 3)随着PEG浓度增大黄花矶松种子萌发率和萌发指数降低, 在-1.5 MPa时无种子萌发; 4)黄花矶松种子萌发率和萌发指数随着NaCl胁迫的加重而逐渐下降, 主要是由渗透胁迫造成, 离子毒害作用甚微; 5)黄花矶松种子适宜播种深度为0.5 cm。
(责任编辑 武艳培)
The authors have declared that no competing interests exist.
| [1] |
|
| [2] |
|
| [3] |
|
| [4] |
|
| [5] |
|
| [6] |
|
| [7] |
|
| [8] |
|
| [9] |
|
| [10] |
|
| [11] |
|
| [12] |
|
| [13] |
|
| [14] |
|
| [15] |
|
| [16] |
|
| [17] |
|
| [18] |
|
| [19] |
|
| [20] |
|
| [21] |
|
| [22] |
|
| [23] |
|
| [24] |
|
| [25] |
|
| [26] |
|
| [27] |
|
| [28] |
|
| [29] |
|
| [30] |
|
| [31] |
|
| [32] |
|
| [33] |
|
| [34] |
|
| [35] |
|
| [36] |
|
| [37] |
|
| [38] |
|
| [39] |
|
| [40] |
|
| [41] |
|
| [42] |
|