{kind=link}
{kind=link}
异色瓢虫对豌豆修尾蚜的捕食功能反应
[杜军利 , 武德功, 黄保宏, 黄伟东, 王增霞]
, 武德功, 黄保宏, 黄伟东, 王增霞]
, 武德功, 黄保宏, 黄伟东, 王增霞]
|
|


第一作者:杜军利(1983-),女,河北武安人,讲师,博士,主要从事作物保护学教学和科研工作。E-mail:[email protected]
为明确捕食性天敌昆虫异色瓢虫( Harmonia axyridis)对豆科植物重大害虫豌豆修尾蚜( Megoura japonica)的猎食能力,本研究在人工气候箱中[温度(24±1)℃,湿度(70%±10%),14 h光照:10 h黑暗(14 h L:10 h D),针对不同生育阶段的异色瓢虫分别设置了7个豌豆修尾蚜的密度梯度进行捕食试验,分析了异色瓢虫对豌豆修尾蚜的捕食功能反应。结果表明,不同生育阶段的异色瓢虫对豌豆修尾蚜的捕食能力差别较大,异色瓢虫雌成虫和4龄幼虫对豌豆修尾蚜的捕食能力较强,日最大捕食量分别为217.39和196.08头·日-1;处理蚜虫的时间较短,分别为0.004 6和0.005 1 d。异色瓢虫各个生育阶段的捕食功能反应类型均属于Holling Ⅱ型圆盘方程;不同生育阶段的异色瓢虫对豌豆修尾蚜的寻找效应随着蚜虫密度的升高而降低。
To clarify the predation abilities and predatory functional response of different stage of Harmonias axyridis on Megoura japonica, an experiment was conducted in an intelligent light indoor incubator operating at a temperature of (24±1) ℃, relative humidity of (70%±10%), and light:dark cycles of 14 h:10 h. There were significant differences between different stages of H. axyridis with respect to their daily predatory capacity on M. japonica. The predation of H. axyridis female adult and fourth instar larvae were stronger than others and their daily maximum prey capacity were 217.39 and 196.08 head·d-1, respecitiverly). They also had a lower duration of possible predation on aphids, which were 0.004 6 and 0.005 1 d, respectively. The functional responses of different stages of H. axyridis to different density of M. japonica fitted well with the Holling Ⅱ model. The searching efficiency of different stages H. axyridis decreased with increasing density of M. japonica.
豌豆修尾蚜(Megoura japonica) 又名蚕豆修尾蚜, 属于半翅目蚜科, 是蚕豆(Vicia faba)、豌豆(Pisum sativum)、大豆(Glycine max) 等豆科作物的重要害虫, 主要在嫩枝和叶片背面为害, 造成茎叶卷缩, 节间缩短, 生长受到抑制, 直接影响豆科作物的产量和品质[1]。化学防治是豆科作物田间防治的常用方法, 但由于化学防治的“ 3R” 问题(抗药性Resistance、再猖獗Resurgence、农药残留Residue) 较为突出[2, 3], 使生物防治成为今后豆科田间综合防治的重要途径。
异色瓢虫 (Harmonia axyridis) 是蚜虫、粉虱、介壳虫等害虫的重要捕食性天敌[4, 5, 6, 7, 8, 9], 其捕食量很大, 对各种蚜虫均具有较强的控制能力。国内外关于异色瓢虫对蚜虫捕食能力及捕食功能反应的研究较多[10, 11, 12, 13, 14] , 如异色瓢虫对大豆蚜 (Aphis glycines)、烟蚜(Myzus persicae)、桃大尾蚜(Hyalopterus arundinis)、葡萄根瘤蚜(Daktulosphaira vitifoliae)、云杉长足大蚜(Cinara alba)等蚜虫捕食功能反应的影响。另外, 温度、群体密度、色型、空间大小等因素对瓢虫的搜索能力均有影响。目前关于豌豆修尾蚜天敌防治的报道较少, 仅见1篇文献报道大草蛉 (Chrysopa pallens) 捕食豌豆修尾蚜[15]。因此, 本研究通过分析不同阶段的异色瓢虫对豌豆修尾蚜捕食功能反应, 以期为豌豆修尾蚜田间综合防治提供理论支撑。
供试虫源异色瓢虫成虫和豌豆修尾蚜均采自安徽省凤阳县安徽科技学院西区种植园, 将异色瓢虫成虫饲养在人工气候室内, 温度设置为(24± 1) ℃, 湿度保持在(70%± 10%), 14 h光照:10 h黑暗, 试验于豌豆修尾蚜盛发期(4-5月)进行, 因此异色瓢虫成虫的饲养也主要用豌豆修尾蚜饲喂, 饲养于塑料盒内(20 cm× 15 cm× 10 cm), 放入几片蚕豆(Vicia faba)叶片作为瓢虫的栖息场所, 也是蚜虫的栖息地, 用医用棉球保湿, 待异色瓢虫成虫产卵后, 把卵转移到新的塑料盒中(条件同成虫), 待卵孵化后用于试验。
待异色瓢虫成虫所产卵孵出后, 即为1龄幼虫, 以后每蜕一次皮增加1龄, 幼虫期总共4龄, 4龄后转化为蛹, 蛹不捕食蚜虫, 蛹过一段时间后转化为成虫。将不同生育阶段的异色瓢虫分别放入培养皿中, 每个培养皿放入1头异色瓢虫, 饥饿24 h, 将不同密度的豌豆修尾蚜与异色瓢虫放入同一培养皿中, 设置7个豌豆修尾蚜的密度梯度[9], 豌豆修尾蚜的密度设置如表1所列, 每个密度分别重复5次。24 h后统计皿内剩余的豌豆修尾蚜数目, 计算平均捕食量。
| 表1 不同豌豆修尾蚜密度梯度 Table 1 Densities of Megoura japonica |
1.3.1 Holling-Ⅱ 圆盘方程
“ 参考武德功等研究的龟纹瓢虫对豌豆蚜捕食功能反应的Holling-Ⅱ 圆盘方程公式[9]
Na= T× a'× N/(1+ a'× Th× N) 。
式中:Na为异色瓢虫捕食豌豆修尾蚜的数量; a'为异色瓢虫的搜寻效率; N为豌豆修尾蚜的密度; T为异色瓢虫总利用时间(因试验时间设为24 h, 故T=1d); Th表示捕食1头豌豆修尾蚜所用时间。将上述方程转化为直线方程:1/Na=1/(T× a'× N) +Th/T, 由于本研究异色瓢虫捕食豌豆修尾蚜的总时间为24 h, 即T=1 d。因此, 方程可转化为1/Na=(1/Na)× (1/N)+Th。
1.3.2 卡方检验 卡方检验公式:
$\chi$2=∑ [(Oi-Ti )2/Ti ] 。
式中:Oi为测定值, Ti为理论数值, 计算出卡方值后检索卡方分布位数表, 最后比较检验两个值之间的相关性。
1.3.3 寻找效应
S=a'/(1+a'ThN)。
式中:a'为异色瓢虫的搜寻效率, N为豌豆修尾蚜的密度, Th表示捕食1头豌豆修尾蚜所用时间。
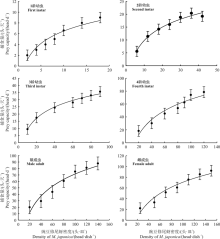
不同阶段异色瓢虫对豌豆修尾蚜的日捕食量随着豌豆修尾蚜密度的升高而增加(表2), 当豌豆修尾蚜达到一定密度时, 异色瓢虫捕食量逐渐趋于平缓, 甚至停止增长(图1)。
| 表2 不同阶段异色瓢虫对豌豆修尾蚜的平均捕食量 Table 2 Average predation of different instars of Harmonia axyridis to Megoura japonica |
异色瓢虫对豌豆修尾蚜的捕食功能反应均属于Holling Ⅱ 型(图1)。所得模型经过卡方检验, 各方程理论值与实测值适合, 所得$\chi$2值均小于(12.592)。
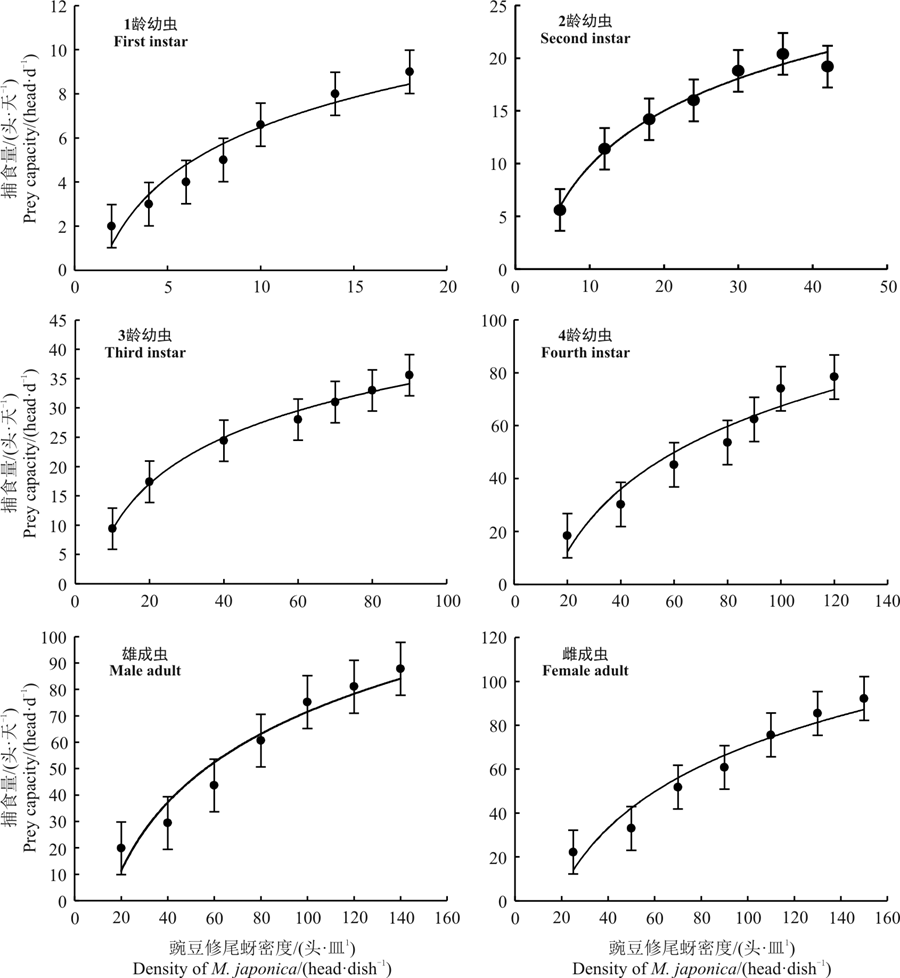 | 图1 不同阶段异色瓢虫对豌豆修尾蚜的捕食功能反应拟合曲线Fig. 1 Fitting curve of the functional response of Harmonias axyridis to Megoura japonica |
异色瓢虫捕食1头豌豆修尾蚜需要0.078 5 d, 即113.040 min; 当豌豆修尾蚜密度较高时, 1龄异色瓢虫对豌豆修尾蚜的最大捕食量为12.74 head· d-1(表3)。2龄幼虫捕食1头豌豆修尾蚜需要0.023 5 d, 即33.840 min, 日最大捕食量为42.55 head· d-1; 3龄幼虫捕食1头豌豆修尾蚜需要0.019 3 d, 即27.792 min, 其日最大捕食量为51.81 head· d-1; 4龄幼虫捕食1头豌豆修尾蚜需要0.005 1 d, 即7.344 min, 日最大捕食量为196.08 head· d-1; 雄成虫捕食1头豌豆修尾蚜需要0.005 5 d, 即7.92 min, 日最大捕食量为181.82 head· d-1; 雌成虫捕食1头豌豆修尾蚜需要0.004 6 d, 即6.624 min, 日最大捕食量为217.39 head· d-1。异色瓢虫的4龄幼虫对豌豆修尾蚜的捕食量明显高于1龄、2龄、3龄和雄成虫, 雌成虫的捕食量最高达到217.39 head· d-1。
| 表3 异色瓢虫对豌豆修尾蚜的捕食功能反应 Table 3 Functional response of different stages of Harmonia axyridis to Megoura japonica |
对实测值和预测值进行$\chi$2检测, 实测值和预测值较为相近(表4)。通过卡方检测发现> $\chi$2, 由此可见, 预测值和实测值无显著差异, 表明异色瓢虫各个阶段的捕食功能反应均属于Holling Ⅱ 型模型, 所以求得的捕食功能反应方程式能够较好地反映出异色瓢虫成虫对豌豆修尾蚜捕食的实际状况。
| 表4 异色瓢虫对豌豆修尾蚜捕食量预测及检验 Table 4 Prediction and test of the feed intake of different instar periods of Harmonia axyridis on Melanaphis sacchari |
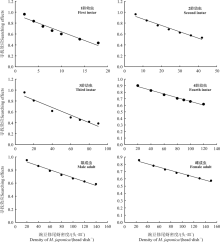
根据方程获得不同豌豆修尾蚜密度条件下不同阶段异色瓢虫与寻找效应的数学模型(表5)。随着豌豆修尾蚜密度的升高异色瓢虫的寻找效应值逐渐降低。随豌豆修尾蚜密度升高, 异色瓢虫1-3龄幼虫的寻找效应值下降幅度较大。1龄幼虫的寻找效应值由0.960 1降为0.435 4, 2龄的寻找效应值由0.960 1降为0.529 8, 3龄幼虫的寻找效应值由0.956 3降为0.386 1; 而4龄及雌雄成虫的寻找效应值降低幅度较小, 其中雌虫的寻找效应值降低幅度最小, 由0.855 9降为0.573 6(图2)。方差分析表明, 3龄幼虫的寻找效应值下降幅度显著高于4龄幼虫(P=0.021 1)和雌成虫(P=0.020 5)。
| 表5 不同豌豆修尾蚜密度与异色瓢虫寻找效应的关系 Table 5 Relationship between density of Megoura japonica and searching effect of Harmonia axyridis |
 | 图2 不同豌豆修尾蚜密度与异色瓢虫寻找效应的关系Fig. 2 Relationship between density of Megoura japonica and searching effect of Harmonia axyridis |
豌豆修尾蚜在安徽省蚌埠地区主要危害蚕豆等豆科作物, 且安徽省蚕豆种植面积较大, 直接影响蚕豆等豆科作物的产量和品质。在每年4-5月为豌豆修尾蚜的盛发期, 此时豌豆修尾蚜的主要天敌有异色瓢虫、多异瓢虫(Adonia variegata)和龟纹瓢虫(Propylaea japonica), 异色瓢虫数量较多, 是豌豆修尾蚜的优势天敌, 在豌豆修尾蚜的自然防控中发挥着重要作用。本研究在室内人工气候箱中研究异色瓢虫不同发育阶段、不同性别对豌豆修尾蚜的捕食情况, 结果显示, 异色瓢虫高龄幼虫和成虫对豌豆修尾蚜有较强的猎食能力, 猎食量随蚜虫数量的增加而增多, 并且各阶段功能反应方程均符合Holling Ⅱ 型方程[16, 17, 18]。异色瓢虫幼虫的捕食量随着阶段的增加而升高, 4龄幼虫对豌豆修尾蚜的捕食量最大, 这与梁洪柱等[19]、林长春等[20]、闫占峰等[21]研究结果一致, 可能是异色瓢虫4龄幼虫通过大量摄取食物蓄积能量为化蛹及羽化为成虫做准备[9, 11]。雌成虫对豌豆修尾蚜的捕食量大于4龄幼虫, 与王东昌等[12]报道的异色瓢虫捕食桃大尾蚜, 林长春等[20]报道的异色瓢虫捕食异毛真胸蚜(Euthoracaphis heterotricha)的研究结果一致; 与梁洪柱等[19]报道的异色瓢虫捕食槐蚜(Aphis sophoricola), 闫占峰等[21]报道的龟纹瓢虫捕食玉米蚜(Rhopalosiphum maidis)研究结果不同, 可能是猎物种类不同导致雌成虫和4龄幼虫猎食偏好不同。这说明异色瓢虫对蚜虫的捕食量与瓢虫所处的生育时期、营养需求及猎物种类有紧密联系。因此, 在田间利用异色瓢虫防治豌豆修尾蚜时, 在蚜虫发生初期释放异色瓢虫4龄幼虫和成虫的防治效果较好。
寻找效应是天敌对猎物猎食的一种行为效应, 天敌对害虫控制作用的强弱与寻找效应呈正相关关系。本研究中寻找效应值随着豌豆修尾蚜密度的升高而逐渐降低, 该研究结果与对异色瓢虫捕食豆蚜[2]和龟纹瓢虫捕食玉米蚜[21]的研究相同。因此, 在田间蚜虫发生初期, 瓢虫数量较少时, 释放异色瓢虫进行防治, 效果会更好。
该研究在人工气候箱中进行, 气候箱恒温恒湿的半封闭系统、试验条件与自然条件具有一定的差别, 由于天敌昆虫在自然界的捕食作用除与种群自身的特点相关外, 还与异色瓢虫的种群密度、自然界的环境因素及空间大小等有关, 因此, 本研究结果可能与自然条件下的捕食量存在一定的区别[22], 但能够为异色瓢虫田间释放及控制豌豆修尾蚜提供一定的参考依据。
(责任编辑 武艳培)
The authors have declared that no competing interests exist.
| [1] |
|
| [2] |
|
| [3] |
|
| [4] |
|
| [5] |
|
| [6] |
|
| [7] |
|
| [8] |
|
| [9] |
|
| [10] |
|
| [11] |
|
| [12] |
|
| [13] |
|
| [14] |
|
| [15] |
|
| [16] |
|
| [17] |
|
| [18] |
|
| [19] |
|
| [20] |
|
| [21] |
|
| [22] |
|